Journey to the Microcosmos produces outstanding videos of microscopic life, and they have featured the volvocine algae before. This new video is entirely focused on the volvocines and on the evolution of multicellularity, and it’s really good.
Journey to the Microcosmos produces outstanding videos of microscopic life, and they have featured the volvocine algae before. This new video is entirely focused on the volvocines and on the evolution of multicellularity, and it’s really good.

I have featured Chytridoodles‘ work before, and I bought a batch of Chlamy print notebooks. Now they have put the same print on face masks. $12.49 for 1, $40 for 4, and shipping is reasonable (if not fast).
![]()
This is straight from the Chlamy 2020 mailing list:
Due to the COVID-19 epidemics, the organizers of Chlamy 2020, the 19th International Conference on the Cell and Molecular Biology of Chlamydomonas, have decided to push all deadlines by 24 days. The early bird registration rates will be available until April 24, i.e. one month before the start of the conference. We hope that this will give you enough time to make an informed decision about your participation to Chlamy 2020. Please visit our website https://chlamy2020.
sciencesconf.org for all details on this top-notch meeting. If you are tempted to participate, please do pre-register now, without paying. It will help us keep track of who would be interested to attend. This way you will receive all the information in due time, and be able to manage travel arrangements so as to benefit from the currently low air fares. Payment of registration fees is suspended for the moment, until we see clearer into our future. But you can deposit your abstract any time. The selection of talks from abstracts is scheduled to take place between April 7 and April 15.
Note that we have pledged to NOT cancel the meeting, whatever happens. At worst, we will postpone it, without changing the location.
Hope to see you at Chlamy 2020 !
The Chlamy 2020 organization committee.
![]()
The 19th International Conference on the Cell and Molecular Biology of Chlamydomonas will be held May 24-29 on Île des Embiez on the French Riviera. The venue looks amazing:

Just in time for holiday shopping, Chytridoodles has a whole line of gear illustrated with a Chlamydomonas print: t-shirts, hoodies, mugs, cell phone cases, and more:


Figure 1 from Salomé and Merchant 2019. Taxonomic Basis of Chlamydomonas and Volvox. Ehrenberg’s drawings of Chlamydomonas and Volvox cells, published in 1838. Cells that belong to the same species are indicated by Roman numerals in the right panel. I, Gonium pectorale; II, Gonium punctatum; III, Gonium tranquillum; IV, Gonium hyalinum; V, Gonium glaucum; VI, Eudorina elegans; VII, Syncrypta volvox; VIII, Sphaerosira volvox; IX, Synura uvella; X, Chlamidomonas pulvisculus; XI, Uroglena volvox. The species was identified as Chlamidomonas pulvisculus but renamed Chlamydomonas reinhardtii in 1888.
In a new(-ish) article in The Plant Cell, Patrice Salomé and Sabeeha Merchant review the history and utility of the green alga Chlamydomonas reinhardtii as a model organism. The article discusses the advantages of Chlamy as a model organism, the scientific questions it has been used to explore, the history of Chlamy research, the characteristics of the species, the existing resources and databases, and genetic and genomic techniques. It’s a good introduction to Chlamy research in a more easily-digestible form than the massive, three-volume Chlamydomonas Sourcebook.

Chlamydomonas colonies from the predation experiment.
I’ll be giving a couple of talks on experimental evolution of multicellularity in the next couple of weeks:
Folks, it’s been fun. I feel like I had a pretty good run as a scientist. I met some amazing people, went to beautiful places, and learned things I never would have imagined (Hodgkinia, WTF?!). With all my frustrations and failures, I’ve never once regretted going back to school and becoming a biologist. But now I need to close the door on all of that and find a new way to make a living.
See, the main project I’ve been working on for the last six years, the one that was supported by a NASA postdoctoral fellowship, and that just came out in Scientific Reports, has been debunked:
My postdoc makes fun of me for having a lousy memory. Not long ago she showed me a paper about microRNAs, and I said I hadn’t read it. She responded, “Yes you have; you blogged about it!” The other day we were discussing the use of antibiotics to prevent bacterial contamination, and I said I thought I might have done that at one time. She told me I had, it was ampicillin, and the concentration.
I’ve been blogging for nearly four years now, and I’ve published well over 400 posts. So I’ve learned that before I sink a bunch of time into writing a new blog post, it’s worth a quick search to make sure I’m not going to repeat myself. When a new paper from Takashi Nakada and colleagues popped up in my Google Scholar alerts, I didn’t immediately realize that I had already written about it. That post was mainly about a new analysis by Thomas Pröschold and colleagues, with the Nakada trees serving as a point of comparison. The new paper is worth its own post, though.
A group of researchers from Keio University have published a new analysis of evolutionary relationships among green algae in the order Volvocales. Takashi Nakada, Yudai Tsuchida, and Masaru Tomita inferred relationships using one nuclear gene and five chloroplast genes.
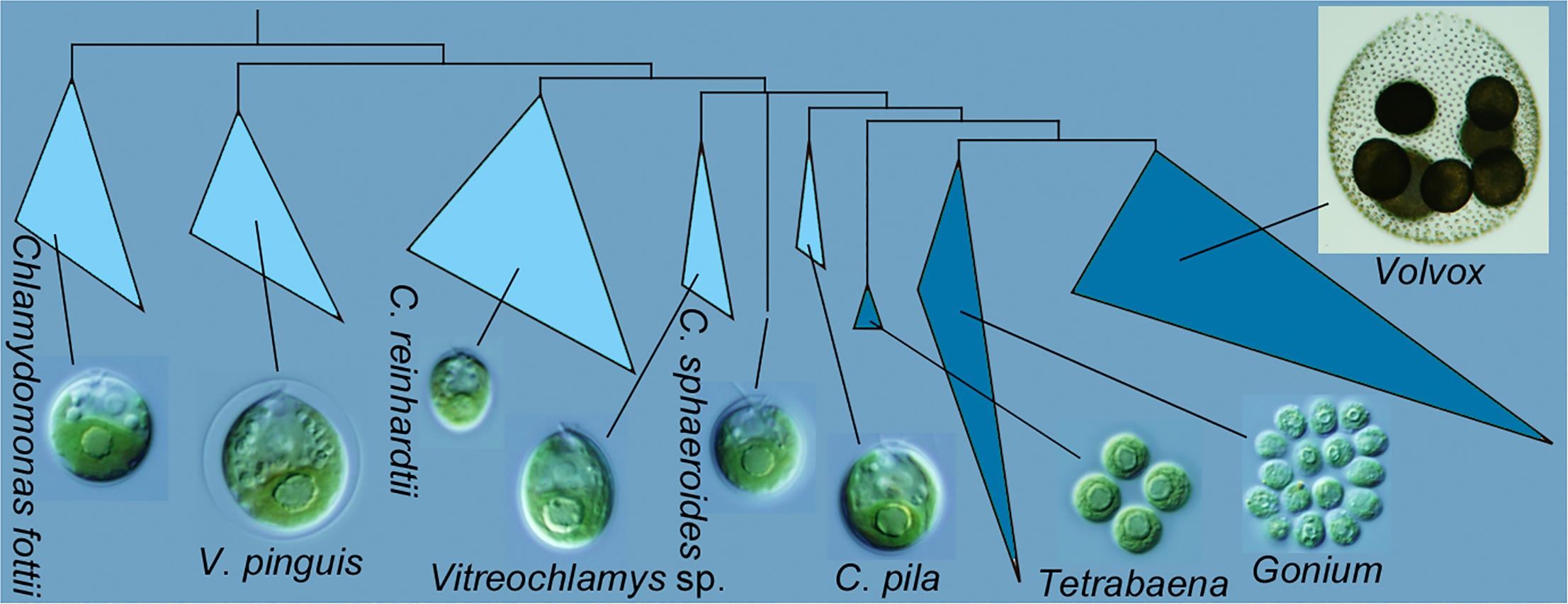
Graphical abstract from Nakada et al. 2019 showing Chlamydomonas pila as sister to the multicellular volvocine algae (Tetrabaena, Gonium, Volvox).
Previously, I focused on the monophyly of the multicellular volvocine algae, i.e. the Tetrabaenaceae, Goniaceae, and Volvocaceae (TGV). The multigene analysis shown above supports monophyly, although the support values for the critical node are not shown (meaning that the Bayesian posterior probability is <0.90 and the bootstrap proportions are <50%). Similarly, the new phylogeny doesn’t do much to resolve the backbone relationships within the Volvocaceae. There are differences from previous analyses that would be important if true, specifically in the positions of Volvox globator (the sole representative of Volvox section Volvox) and of Yamagishiella (which appears as part of an isogamous clade rather than sister to the anisogamous/oogamous Eudorina/Pleodorina/(most) Volvox clade). Neither of these differences is well supported, though, which is typical; most published phylogenies provide poor support for these relationships.

Figure 2 from Nakada et al. 2019. Bayesian phylogenetic tree of core-Reinhardtinia based on combined 18S-atpB-psaA-psaB-psbC-rbcL gene sequences. Corresponding posterior probabilities (≥0.90; left) and bootstrap proportions (≥50%) from maximum likelihood (middle) and neighbor-joining (right) analyses are shown next to the branches. Branch lengths and scale bars represent the expected number of nucleotide substitutions per site. Metaclades (MC; 1.00 posterior probabilities).
The main point of the new paper, though, is the close relationship between the multicellular volvocine algae and Chlamydomonas pila. The critical node for this relationship is is supported by a high Bayesian posterior probability (1.00) but crappy bootstrap values (55% for maximum likelihood and <50% for neighbor joining). The authors did do some analyses with fewer taxa to test this relationship, and those trees did have better support, but they also changed other relationships.
Correctly identifying the closest unicellular relative of the multicellular volvocine algae is critical for reconstructing the first steps in the transition to multicellular life. This is far from the first time that other species of Chlamydomonas and some of Vitreochlamys have been implicated. I’m not aware of any previous phylogeny that includes Chlamydomonas pila, but Chlamydomonas debaryana (for example) is usually closer when it is included.
I wouldn’t say that the evolutionary relationships in this group are fully settled at this point; the particulars vary among authors, depending on the gene(s) analyzed, and even depending on the method of phylogenetic inference. Even the monophyly of the multicellular species has been called into question, though I think it’s definitely too early to be confident in that conclusion. Right now it seems that Chlamydomonas pila is the best contender for the sister species to the multicellular clade, and almost certainly a closer relative to Volvox and co. than Chlamydomonas reinhardtii. As the authors point out, this makes C. pila a good candidate for whole-genome sequencing. The closer a relative to the multicellular group we can find, the better we can resolve which changes are specific to the multicellular clade.
Stable links:
Nakada, T., Tsuchida, Y. & Tomita, M. 2019. Improved taxon sampling and multigene phylogeny of unicellular chlamydomonads closely related to the colonial volvocalean lineage Tetrabaenaceae-Goniaceae-Volvocaceae (Volvocales, Chlorophyceae). Mol. Phylogenet. Evol. 130, 1–8. doi: 10.1016/j.ympev.2018.09.013
Just after my conversation with Jackson Wheat, R. J. Downard invited me to his weekly live stream, Evolution Hour. We talked about the evolution of multicellularity, heritability, and, of course, intelligent design.
