Journey to the Microcosmos produces outstanding videos of microscopic life, and they have featured the volvocine algae before. This new video is entirely focused on the volvocines and on the evolution of multicellularity, and it’s really good.
Journey to the Microcosmos produces outstanding videos of microscopic life, and they have featured the volvocine algae before. This new video is entirely focused on the volvocines and on the evolution of multicellularity, and it’s really good.
I’ve been enjoying the Microcosmos series of videos, including the latest on colonial microbes. There’s a good long section on volvocine algae:
Back when I was a cocky grad student, I wrote a paper that was, in some ways, critical of the work of one of the biggest names in my field. David Kirk, who passed away last year, was among the most important figures in establishing Volvox as a model system for development, genetics, and evolution, among other things. He had published a paper that I thought was unnecessarily progressivist, and I said so in terms that, in retrospect, could have been more diplomatic. In response, Dr. Kirk, whom I had never met, sent me a very thoughtful email thanking me for pointing out some of the problems and politely disagreeing on some other points. Its tone was kind and respectful when annoyed and argumentative would have probably been justified.
In that email, he offered a bet, the stakes of which were to be a beer, that one of the things I had suggested would turn out to be wrong. The issue had to do with inversion, a process that the (mostly) spheroidal algae in the family Volvocaceae undergo during development. I have written about inversion many times on Fierce Roller; in a nutshell, these algae start their lives inside-out, with their flagella on the inside, and invert to get the flagella on the outside, where they can be used for swimming. Their relatives in the genus Gonium also undergo a process of partial inversion, changing from cup-shaped (with the flagella on the concave side) to flat or slightly cup-shaped in the other direction. Dr. Kirk had interpreted Gonium‘s partial inversion as a probable intermediate step that led to the complete inversion characteristic of the Volvocaceae. My reconstructions suggested that incomplete inversion in Gonium had evolved separately from complete inversion in the Volvocaceae, and Dr. Kirk bet me that this would turn out to be wrong.
My postdoc makes fun of me for having a lousy memory. Not long ago she showed me a paper about microRNAs, and I said I hadn’t read it. She responded, “Yes you have; you blogged about it!” The other day we were discussing the use of antibiotics to prevent bacterial contamination, and I said I thought I might have done that at one time. She told me I had, it was ampicillin, and the concentration.
I’ve been blogging for nearly four years now, and I’ve published well over 400 posts. So I’ve learned that before I sink a bunch of time into writing a new blog post, it’s worth a quick search to make sure I’m not going to repeat myself. When a new paper from Takashi Nakada and colleagues popped up in my Google Scholar alerts, I didn’t immediately realize that I had already written about it. That post was mainly about a new analysis by Thomas Pröschold and colleagues, with the Nakada trees serving as a point of comparison. The new paper is worth its own post, though.
A group of researchers from Keio University have published a new analysis of evolutionary relationships among green algae in the order Volvocales. Takashi Nakada, Yudai Tsuchida, and Masaru Tomita inferred relationships using one nuclear gene and five chloroplast genes.
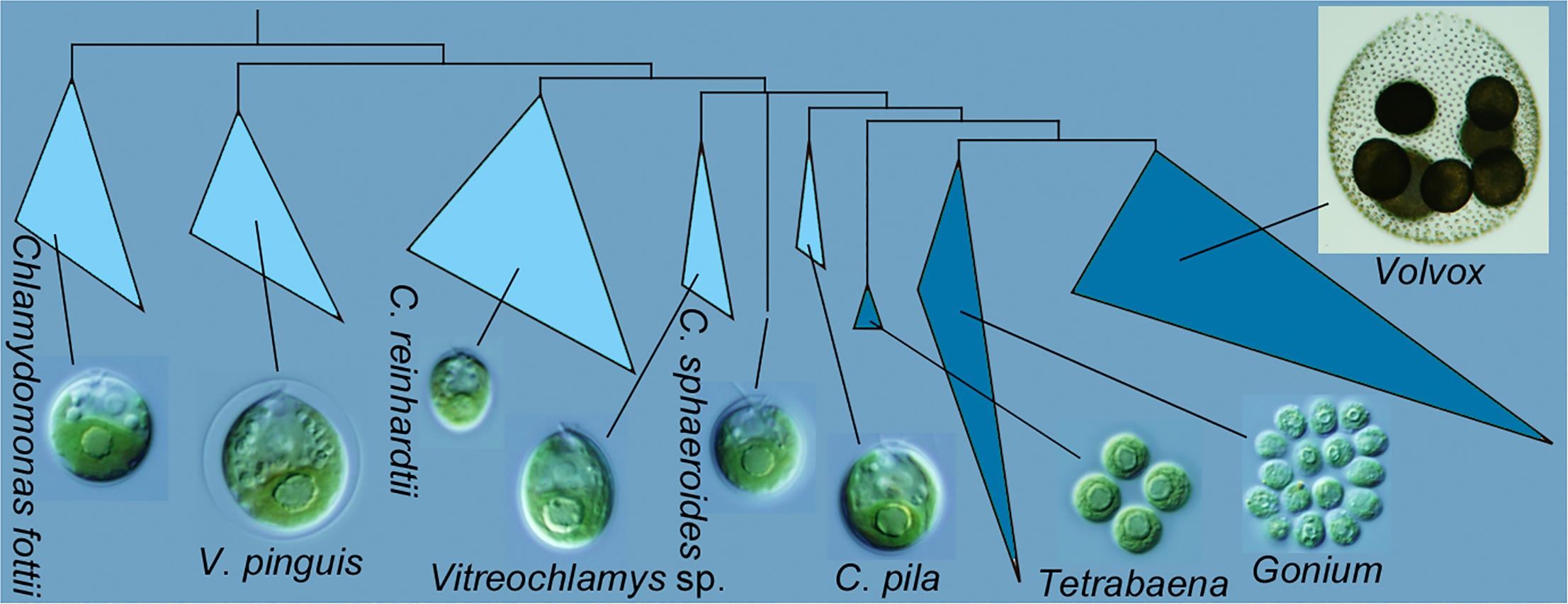
Graphical abstract from Nakada et al. 2019 showing Chlamydomonas pila as sister to the multicellular volvocine algae (Tetrabaena, Gonium, Volvox).
Previously, I focused on the monophyly of the multicellular volvocine algae, i.e. the Tetrabaenaceae, Goniaceae, and Volvocaceae (TGV). The multigene analysis shown above supports monophyly, although the support values for the critical node are not shown (meaning that the Bayesian posterior probability is <0.90 and the bootstrap proportions are <50%). Similarly, the new phylogeny doesn’t do much to resolve the backbone relationships within the Volvocaceae. There are differences from previous analyses that would be important if true, specifically in the positions of Volvox globator (the sole representative of Volvox section Volvox) and of Yamagishiella (which appears as part of an isogamous clade rather than sister to the anisogamous/oogamous Eudorina/Pleodorina/(most) Volvox clade). Neither of these differences is well supported, though, which is typical; most published phylogenies provide poor support for these relationships.

Figure 2 from Nakada et al. 2019. Bayesian phylogenetic tree of core-Reinhardtinia based on combined 18S-atpB-psaA-psaB-psbC-rbcL gene sequences. Corresponding posterior probabilities (≥0.90; left) and bootstrap proportions (≥50%) from maximum likelihood (middle) and neighbor-joining (right) analyses are shown next to the branches. Branch lengths and scale bars represent the expected number of nucleotide substitutions per site. Metaclades (MC; 1.00 posterior probabilities).
The main point of the new paper, though, is the close relationship between the multicellular volvocine algae and Chlamydomonas pila. The critical node for this relationship is is supported by a high Bayesian posterior probability (1.00) but crappy bootstrap values (55% for maximum likelihood and <50% for neighbor joining). The authors did do some analyses with fewer taxa to test this relationship, and those trees did have better support, but they also changed other relationships.
Correctly identifying the closest unicellular relative of the multicellular volvocine algae is critical for reconstructing the first steps in the transition to multicellular life. This is far from the first time that other species of Chlamydomonas and some of Vitreochlamys have been implicated. I’m not aware of any previous phylogeny that includes Chlamydomonas pila, but Chlamydomonas debaryana (for example) is usually closer when it is included.
I wouldn’t say that the evolutionary relationships in this group are fully settled at this point; the particulars vary among authors, depending on the gene(s) analyzed, and even depending on the method of phylogenetic inference. Even the monophyly of the multicellular species has been called into question, though I think it’s definitely too early to be confident in that conclusion. Right now it seems that Chlamydomonas pila is the best contender for the sister species to the multicellular clade, and almost certainly a closer relative to Volvox and co. than Chlamydomonas reinhardtii. As the authors point out, this makes C. pila a good candidate for whole-genome sequencing. The closer a relative to the multicellular group we can find, the better we can resolve which changes are specific to the multicellular clade.
Stable links:
Nakada, T., Tsuchida, Y. & Tomita, M. 2019. Improved taxon sampling and multigene phylogeny of unicellular chlamydomonads closely related to the colonial volvocalean lineage Tetrabaenaceae-Goniaceae-Volvocaceae (Volvocales, Chlorophyceae). Mol. Phylogenet. Evol. 130, 1–8. doi: 10.1016/j.ympev.2018.09.013
Last week, I wrote about Takashi Hamaji’s new paper characterizing the mating-type/sex-determining loci in Eudorina and Yamagishiella. That paper showed that the sex-determining region of anisogamous Eudorina is, surprisingly, considerably smaller than the mating-type loci of isogamous Chlamydomonas, Gonium, or Yamagishiella. Because only one gene, MID, is present in the male version of the sex-determining region in Eudorina, Hamaji and colleagues concluded that
…the evolution of males in volvocine algae might have resulted from altered function of the sex-determining protein MID or its target genes.
I commented that
…we’re still left with two (non-mutually exclusive) possibilities: changes to the MID gene itself may have changed which genes it interacts with (or how it interacts), or there may have been changes in the genes whose expression is controlled by MID.
Now Sa Geng and colleagues have provided at least a partial answer. In a new paper in Development, they swapped versions of MID among different volvocine species* (unfortunately, no unpaywalled version of the paper is currently available; I will add a link when I find one). We already knew that MID is necessary and sufficient for male development: genetically male Volvox carteri colonies that have MID expression turned off produce eggs, and genetically female colonies transformed with MID produce sperm packets (“Sex change (in Volvox)”). But that’s MID from the same species. It’s somewhat surprising that a single gene can cause Volvox to switch sexes, but at least Volvox MID evolved side-by-side with the genes whose expression it controls.
What would be really surprising is if MID from other species, species that diverged from the Volvox lineage ~200 million years ago, worked in Volvox. It would be extraordinarily surprising if MID from a species that doesn’t even have males could control their development in Volvox. It won’t work. Waste of time; don’t bother trying.
Hiroyuki Sekimoto from Japan Women’s University has published a review of sexual reproduction in the volvocine algae and in the Charophyte Closterium in the Journal of Plant Research. In addition to a brief description of the Chlamydomonas sexual cycle, it includes a succinct review of the genetics of sex and sex determination. Unfortunately, the article is paywalled, and my inquiry to the author has so far gone unanswered.

Figure 1 from Sekimoto 2017. The life cycle of Chlamydomonas reinhardtii. Vegetative cells (V) differentiate into mt+ and mt− gametes (G) during nitrogen starvation (−N). Mating types are restricted by mating-type loci (+ and −). When gametes are mixed, the plus and minus agglutinin molecules on their flagellar surfaces adhere to each other, and this adhesion results in increased intracellular cAMP levels. The signal triggers gamete cell wall release and mating-structure activation. Cells then fuse to form binucleate quadriflagellated cells. Zygotes with thick cell walls germinate in response to light and nitrogen supplementation, and undergo meiosis to release four haploid vegetative cells.
Last time, I wrote about Julian Huxley’s 1912 book, The Individual in the Animal Kingdom, and his use of the volvocine algae as an example. I liked most of what he had to say, though I took issue with his assertion that
…all the other members of the family except Volvox…are colonies and nothing more—their members have united together because of certain benefits resulting from mere aggregation, but are not in any way interdependent, so that the wholes are scarcely more than the sum of their parts.
This is, of course, a matter of how we define a multicellular organism, but I think any definition that excludes, for example, Eudorina, is not a very useful one.
This time, I’ll look at the rest of what Huxley had to say about the volvocine algae, most of which is about Volvox:
Julian Huxley was one of the biologists responsible for the merging of Mendelian genetics and Darwinian evolution in the early 20th century, the modern synthesis. His most influential work was Evolution: The Modern Synthesis, published in 1942. Thirty years earlier, though, he published a book on biological individuality, The Individual in the Animal Kingdom. Thankfully, the copyright on this book has expired, so it is now part of the public domain, and a scanned version is available for free in pdf and epub versions from Google.

Any book with Volvox on the cover can’t be all bad!

I asked the lead author of the Gonium genome paper, Erik Hanschen, a few questions, and he kindly agreed to let me to post his responses (see also here, here, and here). I have edited his responses only for formatting:
Blogging took a backseat to the wedding of two dear friends two weekends ago and to morel hunting last weekend, so I’m only now getting around to a post that should have been written weeks ago (I promised on April 22 that it would be out the following week). Last month, Erik Hanschen and colleagues published the Gonium pectorale genome, filling in some crucial bits of the transition to multicellular life in the volvocine algae. This was a big project, taking several years and involving over 20 authors from over a dozen institutions. The final paper is open access in Nature Communications.
I did post an effort to explain some aspects of the paper to the cdesign proponentsists at Evolution News and Views, who, by their own admission, failed to understand it (“After reading this paper, we’re none the wiser.”). I also complained of the science media’s tendency to refer to all algae as ‘pond scum.’ The lead author of the genome paper kindly followed up with a guest post addressing some of ENV‘s other misunderstandings, such as the purpose of model organisms in biology and the difference between ‘assertion’ and ‘evidence’. But now it’s time to dig into what the genome paper actually says.