Journey to the Microcosmos produces outstanding videos of microscopic life, and they have featured the volvocine algae before. This new video is entirely focused on the volvocines and on the evolution of multicellularity, and it’s really good.
Journey to the Microcosmos produces outstanding videos of microscopic life, and they have featured the volvocine algae before. This new video is entirely focused on the volvocines and on the evolution of multicellularity, and it’s really good.
I’ve been enjoying the Microcosmos series of videos, including the latest on colonial microbes. There’s a good long section on volvocine algae:
My postdoc makes fun of me for having a lousy memory. Not long ago she showed me a paper about microRNAs, and I said I hadn’t read it. She responded, “Yes you have; you blogged about it!” The other day we were discussing the use of antibiotics to prevent bacterial contamination, and I said I thought I might have done that at one time. She told me I had, it was ampicillin, and the concentration.
I’ve been blogging for nearly four years now, and I’ve published well over 400 posts. So I’ve learned that before I sink a bunch of time into writing a new blog post, it’s worth a quick search to make sure I’m not going to repeat myself. When a new paper from Takashi Nakada and colleagues popped up in my Google Scholar alerts, I didn’t immediately realize that I had already written about it. That post was mainly about a new analysis by Thomas Pröschold and colleagues, with the Nakada trees serving as a point of comparison. The new paper is worth its own post, though.
A group of researchers from Keio University have published a new analysis of evolutionary relationships among green algae in the order Volvocales. Takashi Nakada, Yudai Tsuchida, and Masaru Tomita inferred relationships using one nuclear gene and five chloroplast genes.
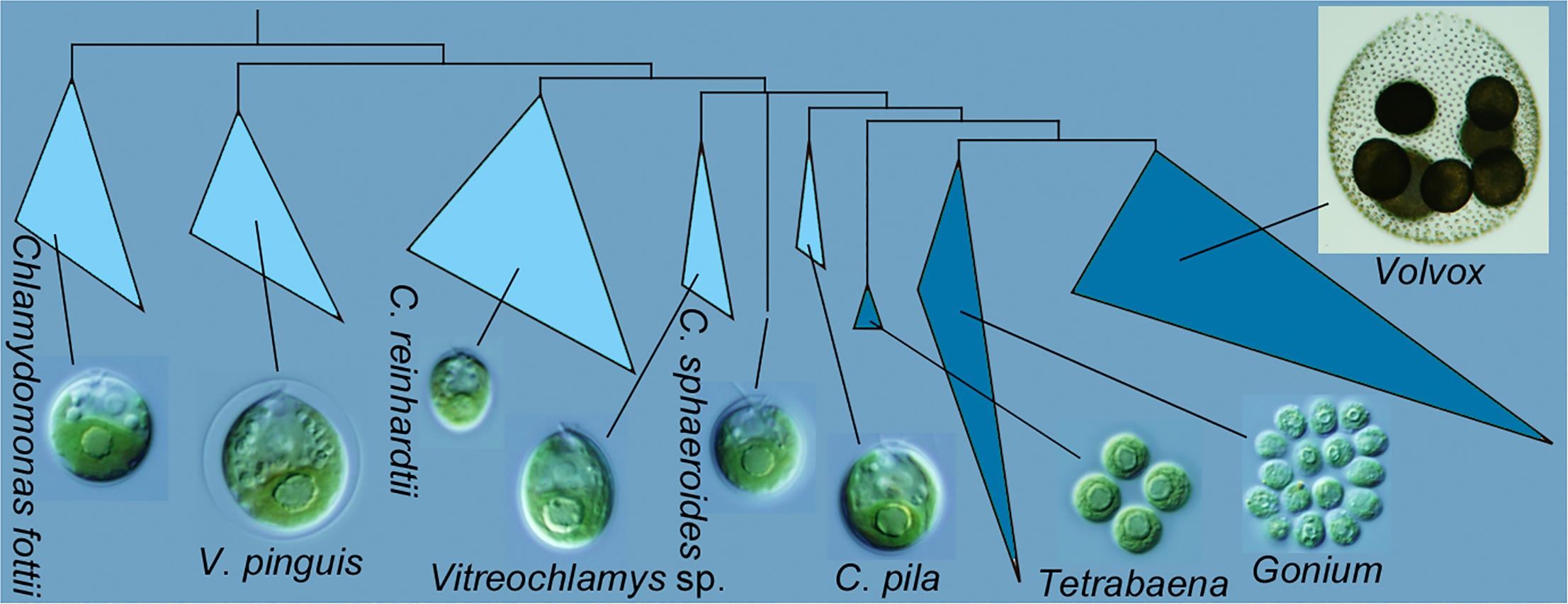
Graphical abstract from Nakada et al. 2019 showing Chlamydomonas pila as sister to the multicellular volvocine algae (Tetrabaena, Gonium, Volvox).
Previously, I focused on the monophyly of the multicellular volvocine algae, i.e. the Tetrabaenaceae, Goniaceae, and Volvocaceae (TGV). The multigene analysis shown above supports monophyly, although the support values for the critical node are not shown (meaning that the Bayesian posterior probability is <0.90 and the bootstrap proportions are <50%). Similarly, the new phylogeny doesn’t do much to resolve the backbone relationships within the Volvocaceae. There are differences from previous analyses that would be important if true, specifically in the positions of Volvox globator (the sole representative of Volvox section Volvox) and of Yamagishiella (which appears as part of an isogamous clade rather than sister to the anisogamous/oogamous Eudorina/Pleodorina/(most) Volvox clade). Neither of these differences is well supported, though, which is typical; most published phylogenies provide poor support for these relationships.

Figure 2 from Nakada et al. 2019. Bayesian phylogenetic tree of core-Reinhardtinia based on combined 18S-atpB-psaA-psaB-psbC-rbcL gene sequences. Corresponding posterior probabilities (≥0.90; left) and bootstrap proportions (≥50%) from maximum likelihood (middle) and neighbor-joining (right) analyses are shown next to the branches. Branch lengths and scale bars represent the expected number of nucleotide substitutions per site. Metaclades (MC; 1.00 posterior probabilities).
The main point of the new paper, though, is the close relationship between the multicellular volvocine algae and Chlamydomonas pila. The critical node for this relationship is is supported by a high Bayesian posterior probability (1.00) but crappy bootstrap values (55% for maximum likelihood and <50% for neighbor joining). The authors did do some analyses with fewer taxa to test this relationship, and those trees did have better support, but they also changed other relationships.
Correctly identifying the closest unicellular relative of the multicellular volvocine algae is critical for reconstructing the first steps in the transition to multicellular life. This is far from the first time that other species of Chlamydomonas and some of Vitreochlamys have been implicated. I’m not aware of any previous phylogeny that includes Chlamydomonas pila, but Chlamydomonas debaryana (for example) is usually closer when it is included.
I wouldn’t say that the evolutionary relationships in this group are fully settled at this point; the particulars vary among authors, depending on the gene(s) analyzed, and even depending on the method of phylogenetic inference. Even the monophyly of the multicellular species has been called into question, though I think it’s definitely too early to be confident in that conclusion. Right now it seems that Chlamydomonas pila is the best contender for the sister species to the multicellular clade, and almost certainly a closer relative to Volvox and co. than Chlamydomonas reinhardtii. As the authors point out, this makes C. pila a good candidate for whole-genome sequencing. The closer a relative to the multicellular group we can find, the better we can resolve which changes are specific to the multicellular clade.
Stable links:
Nakada, T., Tsuchida, Y. & Tomita, M. 2019. Improved taxon sampling and multigene phylogeny of unicellular chlamydomonads closely related to the colonial volvocalean lineage Tetrabaenaceae-Goniaceae-Volvocaceae (Volvocales, Chlorophyceae). Mol. Phylogenet. Evol. 130, 1–8. doi: 10.1016/j.ympev.2018.09.013
Last week, I wrote about Takashi Hamaji’s new paper characterizing the mating-type/sex-determining loci in Eudorina and Yamagishiella. That paper showed that the sex-determining region of anisogamous Eudorina is, surprisingly, considerably smaller than the mating-type loci of isogamous Chlamydomonas, Gonium, or Yamagishiella. Because only one gene, MID, is present in the male version of the sex-determining region in Eudorina, Hamaji and colleagues concluded that
…the evolution of males in volvocine algae might have resulted from altered function of the sex-determining protein MID or its target genes.
I commented that
…we’re still left with two (non-mutually exclusive) possibilities: changes to the MID gene itself may have changed which genes it interacts with (or how it interacts), or there may have been changes in the genes whose expression is controlled by MID.
Now Sa Geng and colleagues have provided at least a partial answer. In a new paper in Development, they swapped versions of MID among different volvocine species* (unfortunately, no unpaywalled version of the paper is currently available; I will add a link when I find one). We already knew that MID is necessary and sufficient for male development: genetically male Volvox carteri colonies that have MID expression turned off produce eggs, and genetically female colonies transformed with MID produce sperm packets (“Sex change (in Volvox)”). But that’s MID from the same species. It’s somewhat surprising that a single gene can cause Volvox to switch sexes, but at least Volvox MID evolved side-by-side with the genes whose expression it controls.
What would be really surprising is if MID from other species, species that diverged from the Volvox lineage ~200 million years ago, worked in Volvox. It would be extraordinarily surprising if MID from a species that doesn’t even have males could control their development in Volvox. It won’t work. Waste of time; don’t bother trying.

Phycologia Volume 56, Issue 4 cover showing Pleodorina sphaerica. Photograph by Hisayoshi Nozaki.
I track the #Volvox hashtag on Twitter, which is how I find out about a lot of the off-label uses of the name Volvox, like DJ Volvox, Volvox the ship, and Volvox the art gallery. Every now and then, it even turns up something related to Volvox the little rolling algae. The other day, @QuintaSwinger tweeted the following video with #Volvox:
The Twitter handle is about just what you think it is; apparently volvocine sex puts someone in mind of polyamory. I suppose I can see that: when a sperm packet enters a colony, it gets busy with all the ova. The video was uploaded to YouTube by Dr. Donald Ott from the University of Akron.
I think the algae in the video are not actually Volvox, though. Certainly the still photo at the beginning is Volvox. Probably not section Volvox (too few cells), and probably not Developmental Program 2 (germ cells too small in the one on the lower right). If I had to guess, I’d say V. aureus, but that’s largely a Bayesian bet because they’re so common. Maybe Alexey Desnitskiy or Hisayoshi Nozaki can comment.
The colonies in the video, though, look more like Pleodorina to me. Not P. sphaerica, since the somatic cells are all in the front, but without more information I can’t narrow it down more than that.

Figure 1 from Nozaki et al. 2017. Pleodorina sphaerica.
There really aren’t enough people looking for volvocine algae. There’s a suspicious tendency for the geographical centers of volvocine diversity — southern Africa, central North America, southeast Asia — to include the home institutions of phycologists studying volvocine diversity — Mary Pocock, Richard Starr, Hisayoshi Nozaki, respectively. I find it much more likely that this is an artifact of sampling effort than that, for example, central Africa and Central and South America are depauperate of volvocine algae.

Figure 3 B-I from Knysak and Żelazna-Wieczorek 2017. Pleodorina indica (Iyengar) H. Nozaki 400x.
A new paper in Oceanological and Hydrobiological Studies reports a massive bloom of Pleodorina indica in a reservoir in central Poland. Piotr Knysak and Joanna Żelazna-Wieczorek sampled the reservoir on the Olechówka River in Łódź during the summer of 2015 and found that P. indica made up ~95% of the algae collected.
Stephanie Höhn and Armin Hallmann have published a detailed study of the developmental process of inversion in Pleodorina californica. Pleodorina is one of the two genera we usually refer to as ‘partially differentiated’ (the other is Astrephomene), meaning that some of their cells are specialized for motility and never reproduce (soma) and some perform both motility and reproductive functions. P. californica is pretty big, up to about 1/3 of a millimeter, easily visible to the naked eye (though you’d need better vision than mine to make out any details).

Stephanie Höhn sampling a pond near Cambridge University during the Volvox 2015 meeting.
Like all members of the family Volvocaceae, P. californica undergoes complete inversion during development:
After the completion of the cell division phase and before inversion, the embryos of Gonium, Pandorina, Eudorina and Pleodorina consist of a bowl-shaped cell sheet, whereas the embryonic cells of Volvox form a spherical cell sheet. With exception of the genus Astrephomene, all multicellular volvocine embryos face the same “problem”: the flagellar ends of all the cells point toward the interior of the bowl-shaped or spherical cell sheet rather than to the exterior, where they need to be later to function during locomotion. [References removed]

32- and 64-celled colonies of Pleodorina starrii. Not to scale. 
Pleodorina starrii by Matthew Herron is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License.
I spent a year in graduate school trying to cross male and female strains of the volvocine green alga Pleodorina californica. A year. I did some other stuff in that time, but I spent an awful lot of it trying to convince these algae to get busy. I threw everything I could think of at them: four different mating media, different temperatures, different lighting conditions…nothing worked. I never recovered a single viable zygote. I needed to cross them to generate some genetic variation for an ambitious artificial selection study, my ‘official’ dissertation project. Eventually, my advisor suggested I ask Hisayoshi Nozaki for advice.
There is little doubt that Dr. Nozaki is the world’s leading expert on volvocine biodiversity, having described about half of the known species (see for example New Volvox Species, Volvox ovalis, and African Volvox in Montana). He responded that the strains of Pleodorina californica I had been failing to breed had been collected many years ago and had probably lost the ability to reproduce sexually (a problem I mentioned in Why don’t we revise volvocine taxonomy?). I had been spinning my wheels, never realizing that I had no hope of success. I should have contacted Dr. Nozaki about eleven months earlier.