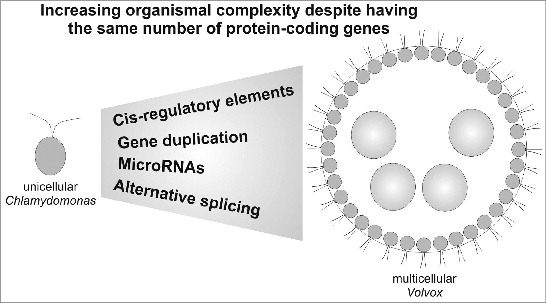
Figure 1 from Kianianmomeni 2015. Gene regulatory mechanisms behind the evolution of multicellularity. Model illustrating the role of gene regulatory mechanisms in the evolution of multicellular Volvox from a Chlamydomonas-like ancestor.
Arash Kianianmomeni’s latest paper in Communicative & Integrative Biology addresses the possible roles of gene regulation and alternative splicing in the evolution of multicellularity and cellular differentiation (Kianianmomeni, A. 2015. Potential impact of gene regulatory mechanisms on the evolution of multicellularity in the volvocine algae. Commun. Integr. Biol., 37–41. doi 10.1080/19420889.2015.1017175). The article is an ‘Addendum’ to a 2014 study by Kianianmomeni and colleagues in BMC Genomics. Communicative & Integrative Biology often invites authors to write these addenda after they have published a (usually high impact) paper elsewhere, providing authors the opportunity to publish material that was not included in the original paper due to space limitations or because it was opinionated or speculative. I may address the BMC Genomics article in a future post, but right now there is more new volvocine research than I have time to write about (it should be an exciting Volvox meeting this summer!).
The main point of the new article is that the transition to multicellular life, at least in the volvocine algae, is more likely due to changes in gene expression than to changes in gene content. The nearly identical gene content of the Chlamydomonas reinhardtii and Volvox carteri genomes leads Kianianmomeni to conclude that
…the transition from a unicellular Chlamydomonas-like ancestor to multicellular Volvox did not require major changes in gene content.
I made a similar argument in response to Ann Gauger’s bizarre assertions to the contrary:
Neither microtubules or [sic] kinesin are [sic] present in bacteria so their origin must be accounted for…This process [apoptosis] involves a minimum of several novel genes as well…Getting two kinds of reproductive cells, eggs and sperm, requires triggering a new set of regulatory genes…[emphasis mine]
Kianianmomeni’s view is in direct opposition to Dr. Gauger’s:
…the development of species-specific proteins could not account for the development of Volvox from a Chlamydomonas-like ancestor.
I’m not sure what led Dr. Gauger to conclude that each new trait requires the origin of one or more genes, but Kianianmomeni’s conclusions are based on, you know, evidence:
This is also supported by experimental evidence that showed that 2 important proteins, GlsA and InvA, which are responsible for essential developmental processes behind the evolution of multicellularity in Volvox, namely asymmetric division and embryo inversion, respectively, are conserved in unicellular Chlamydomonas. Interestingly, Chlamydomonas orthologs can rescue Volvox glsA and invA mutants… We and others have recently shown that alternative splicing could contribute to the appearance of multicellularity by generating multiple transcripts from a single gene.
Kianianmomeni goes on to suggest that alternative splicing could be an important factor in the evolution of multicellularity, and that a genome-wide comparison of alternative splicing in Chlamydomonas and Volvox would test this prediction. After reviewing the roles of cis-regulatory elements, gene duplications, and micro-RNAs in Volvox development, he concludes that
The evolution of gene regulatory mechanisms, rather than the development of species-specific proteins, seems to play a central role in diverse developmental processes during the unicellular-multicellular transition.
I don’t think this conclusion will be controversial; after all, we have known for some time that important innovations in the Volvox lineage, mostly related to cellular differentiation, were related to changes in gene expression. There is good reason to think, though, that at least one important change, the increased size and structural complexity of the extracellular matrix (ECM) in Volvox compared to Chlamydomonas, involved gene duplications. Two of the very few differences in gene content between the two genomes were expansions of gene families involved in building the ECM: the Volvox matrix metalloproteases and the pherophorins.
I’d agree that the control of how genes function in the Volvocines is more likely to be the primary mechanistic force at play. Maybe this is true for most major developments in evolutionary biology. But I have to add some caveats. It’s being a nit-picking but honestly I don’t think we know the function (let’s say major function) of half of the Volvocine genes. Only about half (about 8000) of Volvox proteins are easily recognisable as being reciprocal hits with Chlamy proteins. That’s some pretty big divergence. The number of genes in both organisms is also largely a function of the annotation process and in both organisms there are some big issues with annotation. The latest versions of Chlamy are good but the new version of Volvox really isn’t. If memory serves only about 60% of Volvox V1 genes had protein or est evidence. The rest were in silico predicted and again for both organisms there are thousands of hypothetical proteins that perhaps possess a domain but other than that we can’t ascribe them a function. Domain scans after all are based on what is in our databases. These black box proteins probably remain somewhat as unknown unknowns. Who knows if any have some importance towards multicellularity? Your own work showed us that this transition happened a lot earlier than we had thought. For now we still focus on the genes/proteins we know from earlier work but perhaps there are many differences amongst genes that we’re not familiar with. We still haven’t identified the lag genes. Volvox has another protein very closely related to GlsA and a bunch of extra kinesins. I think we are being premature all saying that the protein-content between these organisms is very close. But just my 2c.
Still, I do think that Kianianmomeni is most likely right. The splicing aspect is also very interesting and makes me ponder about the role of intron size expansion in relation to splicing.
Sorry spamming here…. But what is pretty cool is just how easy it is to test things now like the relative rates of alternative splicing. Not only between species but even between germ-soma.