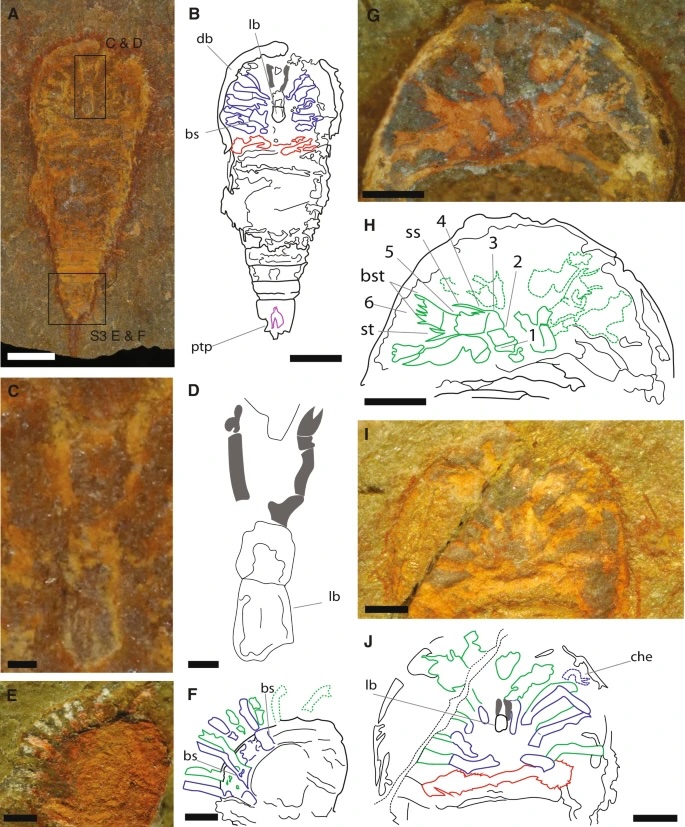
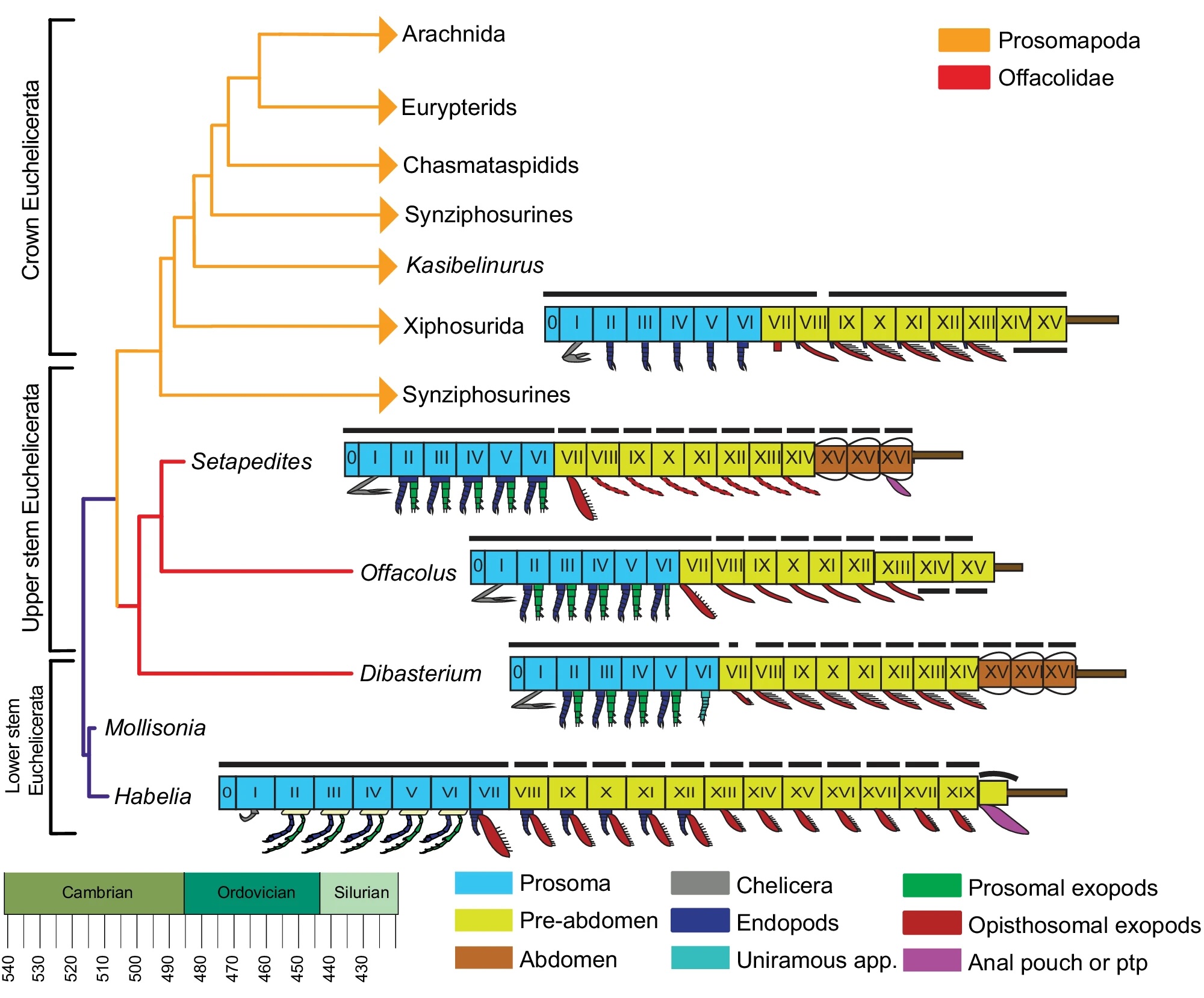
If you don’t care about birds flitting about, here’s another tool, TimeTree. It’ll let you look up the divergence time between any two species, in this case I just chose to compare myself with my house spiders.
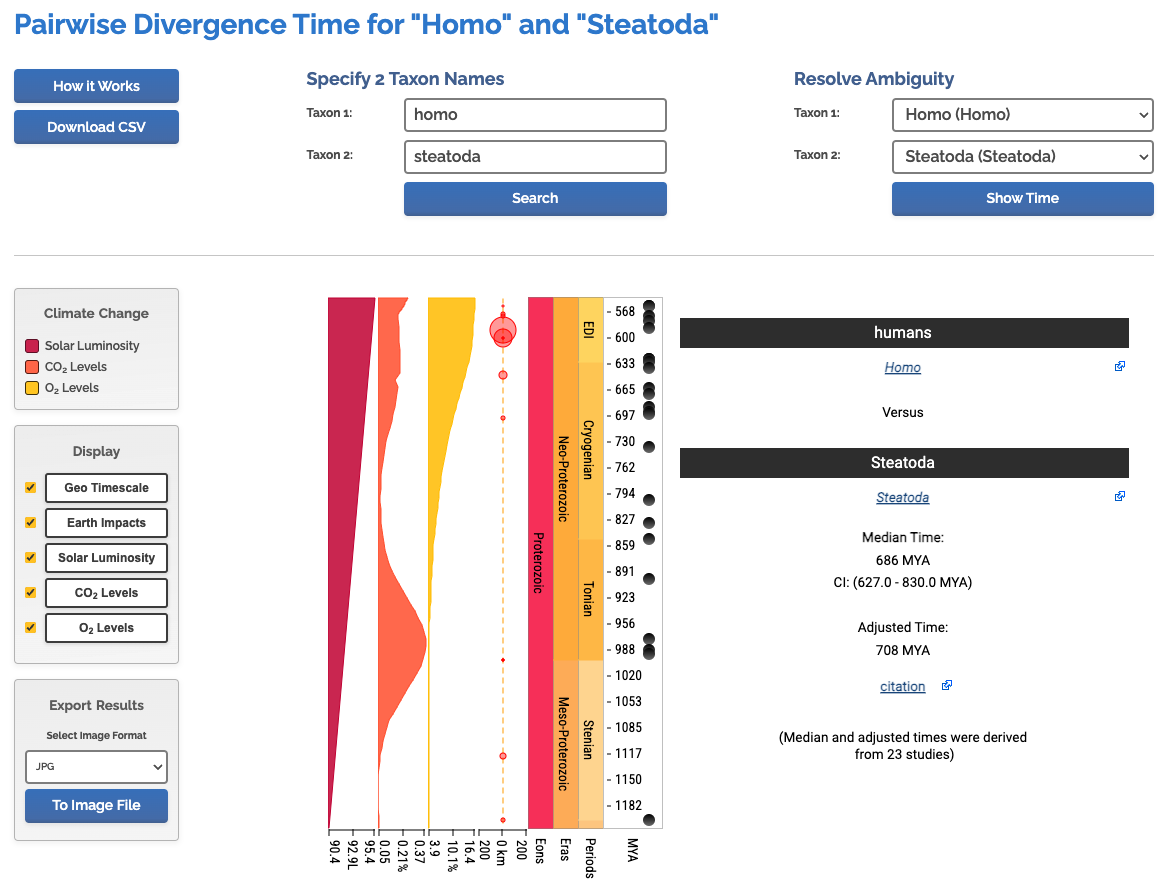
700 million years sounds about right, but that’s just the general time since the last common ancestor of vertebrates and invertebrates. Our shared ancestor would have been some nondescript little worm.
If you’re arguing with Kent Hovind, it might be useful to know that the last common ancestor of humans and bananas lived about 1.5 billion years ago; same time since we diverged from the amoeba. The paleo-proterozoic was a busy time! Or, at least the seas were full of eukaryotes then.