My students are also blogging here:
A good portion of what I’ve been teaching so far uses Drosophila as a model system — it’s the baseline for modern molecular genetics. Unfortunately, it’s also a really weird animal: highly derived, specialized for rapid, robust development, and as we’ve learned more about it, it seems it has been layering on more and more levels of control of patterning. The ancestral system of establishing the body plan was far simpler, and evolution has worked in its clumsy, chance-driven way to pile up and repurpose molecular patterning mechanisms to reinforce the reliability of development. So I promised the students that this would be the last day I talk about insects for a while…we’ll switch to vertebrates so they can get a better picture of a simpler, primitive system. What we’ll see is many familiar genes from flies, used in some different (but related!) ways in vertebrates.
But today I continued the theme of epistatic interactions from last week. Previously, we’d talked about gap genes — genes that were expressed in a handful of broad stripes in the early embryo, and which were regulated in part by the even broader gradient of bicoid expression. The next level of the hierarchy are the pair rule genes, which are expressed in alternating stripes — 7 pairs of stripes for 14 segments.
First point: notice that we are seeing a hierarchy, a descending pattern of regulatory control, and that the outcome of the hierarchy is increasing complexity. One gene, bicoid sets up a gradient that allows cells to sense position by reading the concentration of the gene; the next step leverages that gradient to create multiple broad domains; and the pair rule genes read concentrations of gap genes and uses the boundaries between them to set up even more, smaller and more precise domains of stripes that establish the animal’s segments.
This is epigenesis made obvious. The 14 stripes of the pair rule genes are not present in the oocyte; they emerge via patterns of interactions between cells and genes. The information present in the embryo, as measured by the precise and reproducible arrays of cells expressing specific genes, increases over time.
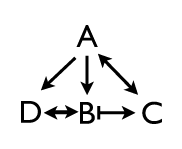
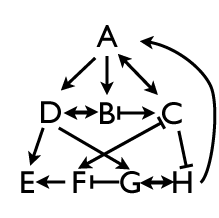
So part of the story is hierarchy, where a complex pattern at one stage is dependent on its antecedents. But another part of the story is peer interaction. Cells are inheriting potentials that are established by a cascading sequence of regulatory events, but in addition, genes at the same approximate level of the hierarchy are repressing and activating each other. We can tease those interactions apart by fairly straightforward experiments in which we knock out individual pair rule genes and ask what the effect of the loss has on other pair rule genes. I led the students through a series of epistatic experiments which started out fairly easy. Knock out a pair rule gene that is expressed in odd numbered parasegments, for instance, and it’s complement, the pair rule gene expressed in even parasegments, expands its expression pattern to fill all segments. Sometimes.
Some of the experiments reveal simple relationships: hairy suppresses runt, and runt suppresses hairy. That makes sense. They have mutually exclusive domains, so it’s no surprise that they exclude each other. But then we looked at other pair rule genes which are expressed in patterns slightly out of phase from the hairy/runt pair, and there the relationships start getting complex. Genes like fushi tarazu are downstream from all the others, and their effects are straightforward (their loss doesn’t disrupt the other pair rule genes), but genes like even-skipped have much messier relationships, and the class was stumped to explain the results we get with that deletion.
So I asked them to come up with other experiments to tease apart these interactions. I was somewhat amused: when I think along those lines, I come up with more genetic crosses and analyses of expression patterns — I think about regulatory logic and inferring rules from modifications of the pattern. Students nowadays…they’re so much more direct. They want to go straight to the molecular biology, taking apart the genes, identifying control elements, building reporter constructs to see gene-by-gene effects. I felt so old-fashioned. But we also had to talk about the difficulty of those kinds of experiments, and that often the genetic approach is better for building a general hypothesis that can be fruitfully tested with the molecular approach.
Then we stopped — we’ll come back to flies later, and start looking at some specific subsets of developmental programs. Next, though, we’re going to take a big step backward and look at early events in vertebrates and progress through that phylum until we see how they build segments. I’m hoping the students will see the similarities and differences.