I could not leave the scarring of this lovely video unaddressed, so I made a video of my own.
Transcript below.
I could not leave the scarring of this lovely video unaddressed, so I made a video of my own.
Transcript below.


Well, sort of. Researchers in the College of Biological Sciences on the Twin Cities campus have created a synthetic cell from scratch that has many of the properties of evolved cells, but they also have significant limitations. Kate Adamala and Aaron Engelhart have combined 7 plasmids in a lipid bubble, that they’ve called a spudcell, that can do many of the cool things that natural cells can do.
This is likely the most exciting project I’ve ever worked on,” said Adamala. “We’ve replicated in chemistry what only used to be possible in biology: the complete set of behaviors of a cell. It proves that the most fundamental functions of life, like growth and replication, do not need a mysterious magical spark.
You can almost hear the mad cackle. So what can it do?
Among the characteristics of SpudCell:
• Replicates a biological cell’s life cycle: SpudCell is capable of selection, genome replication, growth, resource acquisition via feeding, and genetically encoded division.
• Cell division without a cytoskeleton: Natural cells divide using internal scaffolding called a cytoskeleton, which has been a bottleneck in synthetic cell research. SpudCell sidesteps the need for a cytoskeleton with proteins that crowd together on the membrane surface until the mechanical stress makes the membrane split.
• Selection and competition: Researchers introduced a genetic change that increased production of the fusion protein, resulting in cells that grew faster and produced more offspring. After five generations, the faster-growing variant had outcompeted the original. Under nutrient scarcity, the advantage increased, demonstrating selection and competition operating in a fully synthetic chemical system.
It’s basically a minimal, defined biochemical system in a liposome, with a tiny genome. It doesn’t have any organelles (other than ribosomes), and is a uniform soup of enzymes, which is why I think they called it a SpudCell, because it’s got all the character of a potato. You can read a preprint which gives all the details.
The system contains 36 purified enzymes, a 90,000 base pair genome spread across nine separate DNA molecules, and a lipid membrane. SpudCell is able to grow, replicate its genome, divide, and undergo selection and competition across multiple generations.
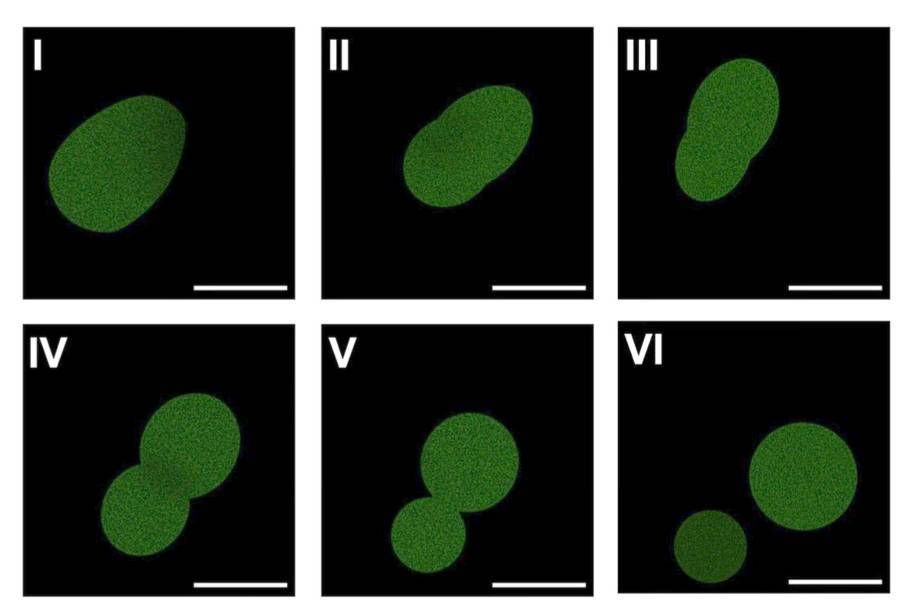
The preprint also discusses some of the major limitations of this system. It’s not going to compete with natural cells, and it will die out within a few generations if released. It needs to be fed specially prepared liposomes filled with nutrients that can spontaneously fuse with the cells, which is kind of cheating. But also…
• Building ribosomes from genetic instructions. SpudCell currently uses ribosomes from E. coli bacteria. Without the capability to remake ribosomes, SpudCell runs for 5-10 generations before the machinery degrades. Building ribosomes from scratch means synthesising dozens of proteins and RNA molecules, then getting them to assemble in the right order.
• Improving genome distribution. After five generations, about 30% of daughter cells have the complete set of seven DNA plasmids. Natural cells solve this with cytoskeletal machinery that pulls chromosomes apart during division. SpudCell does not have that yet, and better genome inheritance will need more sophisticated division mechanisms.
• Reducing dependence on external feeding. Nutrient-carrying liposomes have to be added regularly, and division requires streptavidin and molecular linker proteins from outside. Making the system more autonomous will require building metabolic pathways that can synthesize components from simpler starting materials.
It’s not sophisticated enough to assemble its own ribosomes, which is a major limitation. It’s going to need at least an order of magnitude more complexity to do that.
I’m more interested in the lack of a cytoskeleton. You’ve got 23 pairs of chromosomes in your cells, and the cytoskeletal machinery in mitosis works to guarantee that each daughter cell inherits the same 23 pairs; there are cells that don’t have that cytoskeleton, and they can do just fine with a single strand of circular DNA, but these SpudCells have at least 7 separate plasmids that have to be sorted. They rely on chance distribution of the plasmids to daughter cells, so they have to point out that after 5 generations 70% are nonviable.
What it’s good for is that it’s greatly reduced, bare-bones cell where every component is precisely known so you can track every molecule for at least a few generations. There are toy cells, but good educational toys.

As an undergraduate, my introductory neuroscience course was taught by Johnny Palka, a developmental biologist and neuroscience who worked with Drosophila, who had to explain to us on the first day of class that flies have brains. It was memorable because I was surprised that anyone thought otherwise (don’t worry, the class got much more sophisticated and mathematical after that). But it’s true that there are an awful lot of people with that degree of ignorance.
“Most people think insects are reflex-based machines,” said Dr Olli Loukola, a behavioural ecologist at the University of Oulu, Finland, and senior author. “That they can’t have any emotional states or feel pain. Some people don’t even realise that they have brains. I hope that these results change the worldview about that.”
That’s from an article about bee intelligence, and if you think insect anatomy is a confusing topic for the general public, wait until you find out there are people who think intelligence can be reduced to a single number.
Only…biology can surprise you. We don’t even understand what intelligence is, so you should avoid limiting preconceptions. All it takes is a simple test to demonstrate the capabilities of insects.
The bees, which were only a couple of weeks old, were first trained to associate a blue artificial flower with a reward of sugar water. During the test, the flower was moved to the ceiling of a transparent petri dish-style chamber whose ceiling was too high for them to reach, but with insufficient space for them to hover. A ball was also introduced into the chamber. To reach the flower, the bee had to roll the ball under it and climb on top – a behavioural sequence they had never previously encountered or been trained to perform.
In the most basic version of the test, 75% of the bees were successful in reaching the flower. “This is essentially an insect version of the classic ‘box-and-banana’ problem,” said Loukola. “The animal must realise that an object can be repositioned and then used as a tool to reach an otherwise inaccessible goal. What stands out about the result is that this kind of spontaneous problem solving is now demonstrated in an insect.”
This is not to say that bees have the breadth of ability that a chimpanzee has. It’s saying that some things we regard as a significant intellectual capability can be implemented with a tiny number of neurons, and that includes tool use.
“There’s a general perception that intelligent behaviour requires big brains because we are big-brained and relatively intelligent among animals,” Chittka added. “Bees are a model of how much intelligence you can squeeze into a small nervous system … It’s a good reminder of there being a motivation to pay some respect to these other beings.”
Another example I’d point to is corvids — teeny tiny little bird brains that are remarkably smart.

My daughter won first prize in a poster session at UW Madison, presenting her work on “Evaluating Retrieval-Augmented Generation vs. Long-Context Input for Antibiotic Timeline Extraction from EHRs”. I struggled to follow it, but got the gist of it — they’re working on methods to more efficiently extract information from patients’ medical records using LLMs. She sent us the poster image, maybe you can extract more details from it.
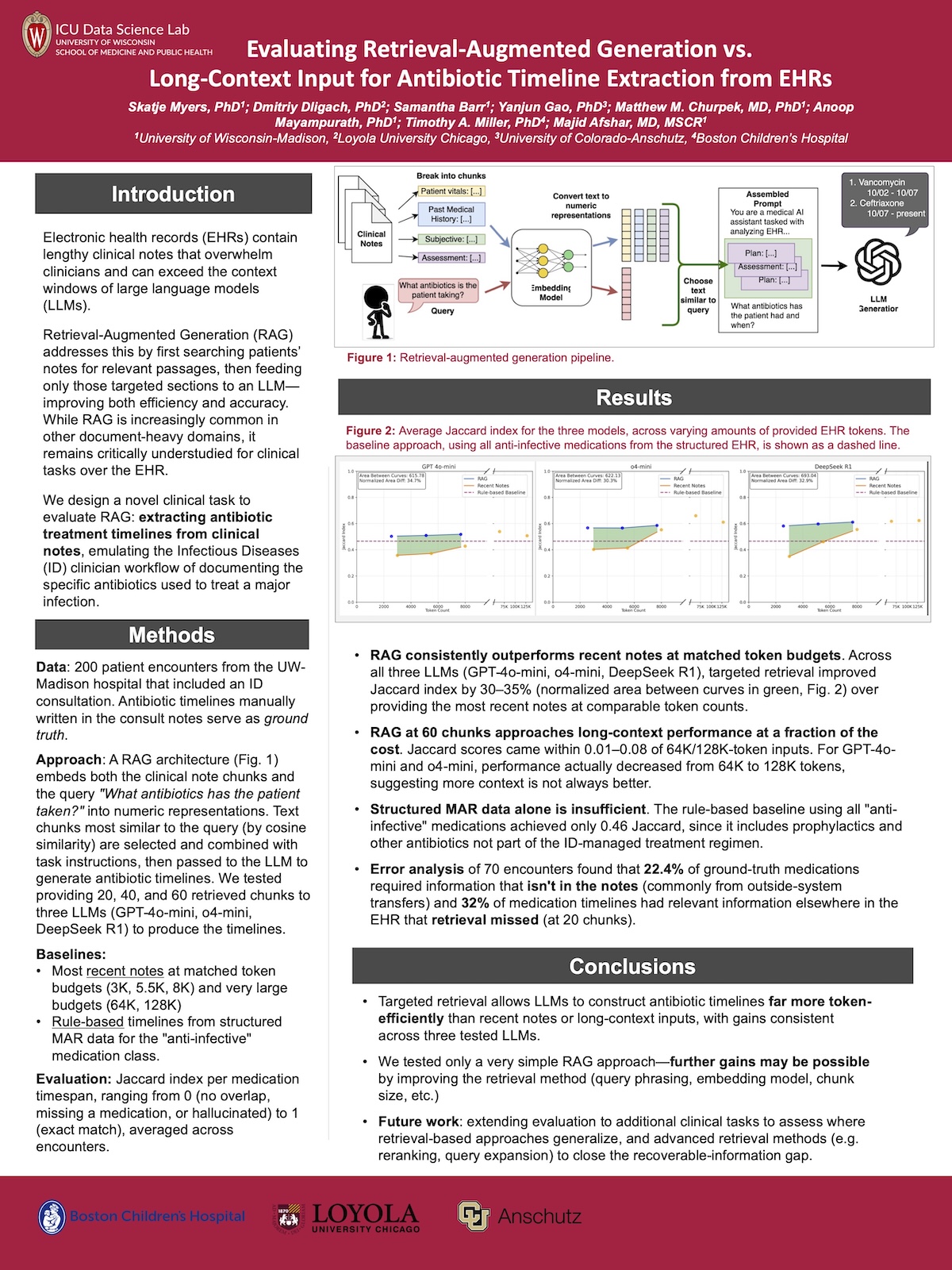
Near as I can tell, it’s perfect, and her peers also thought so. The only suggestion I could possibly make is to maybe add a few spider photos…or a picture of my granddaughter? I don’t know that my suggestions would necessarily help.
It’s nice to have a vague idea of what she’s been up to!

An article published in the journal Diabetes Care, a journal of the American Diabetes Association, and authored by credible scientists in the field, begins with this condemnation of the policies of the NIH administration:
Just a year ago, in these very pages, we highlighted the many threats the current U.S. administration posed to the health of our nation. Since then, there have been actions by the administration that have caused grave health consequences, and their current approach will continue to do so. The numerous measles outbreaks and associated avoidable deaths have resulted in part from hyping disproven theories of harm rather than publicizing the effectiveness of the measles vaccine. Plugging the concept that diabetes is curable by “changing the food source” simply ignores the large body of work that has demonstrated that it is not merely a disease of poor nutrition and the immense challenges of reinventing the food industry. Peddling conspiracy theories represents failures by officials of the Department of Health and Human Services (HHS), whose primary goal is to protect our health. These two examples represent just two of the broken promises made by the current HHS leadership during their confirmation hearings. And, despite promising oversight, representatives on Capitol Hill have shirked their responsibility and have allowed the country to continue along misguided paths that even they recognized as irresponsible.
It goes on at considerable length, documenting all the failures of Jay Bhattacharya and RFK jr. Read the whole thing.
The authors were at a meeting of the American Diabetes Association handing out copies of that editorial to attendees, when the society administration called in uniformed security to kick them out of the meeting, and cancelled their registration.
The incident took place Friday morning at a meeting of the American Diabetes Association in New Orleans, shortly before Dr. Jay Bhattacharya, director of the National Institutes of Health, was scheduled to speak. An organizer announced just before Dr. Bhattacharya’s session that he would no longer be speaking; a senior adviser at the N.I.H. took his place.
The researchers were handing out copies of the editorial, recently published in the association’s flagship journal, which detailed the effects of N.I.H. cuts and other Trump administration actions on diabetes research and outcomes, when security staff asked them to step outside and tried to take away the papers, said Aaron Kelly, a professor of pediatrics at the University of Minnesota who was among the researchers escorted out. A video taken by MedPage Today, which first reported the news, shows a tense confrontation, including a man in uniform putting his hands on an expert.
Wow. That’s fascism in action, suppressing the free discussion of scientific assessment of the field. And this action was ordered by the officers of the American Diabetes Association!
Hours after he was removed, Dr. Kahn received an email from the American Diabetes Association stating that his behavior had violated the conference’s code of conduct.
“You were respectfully informed that distribution of materials was not permitted and given the opportunity to remain in the meeting if you stopped handing out the materials,” the email read. “When you continued the behavior, we had no choice but to remove you for the remainder of the meeting.”
The email went on to note that there are “a lot of logistics and security measures taken when a federal official is in attendance.” A spokesperson for the N.I.H. did not immediately respond to a request for comment.
Several of the researchers said they believed they were removed partly because the association feared repercussions from the Trump administration.
The order refusing to allow the distribution of papers is pure bullshit. It is common practice for researchers to bring along copies of their relevant papers and hand them out. I’ve been at poster sessions where the author brings in a small stand and has a stack of papers that people can take away. Why would you limit the dissemination of information at a meeting whose purpose is to disseminate information?
That it was a “security measure” is equally disingenuous. What was the guy going to do, rush the stage with his papers and slash Bhattacharya with vicious paper cuts?
This action by the American Diabetes Association was simply cowardice by the organizers of the conference. They should be ashamed. This is going to go in the history books as another example among many of chickenshit institutions caving to their own oppression by the Trump administration.
Dr. Kahn said that the editorial aimed to advance science.
“It is no longer enough to stand idly by or work behind the scenes with lawmakers,” the experts wrote in their editorial. “Moreover, it is no longer appropriate to fret about political backlash. Now is the time to recognize and fight to reverse the spiraling fall of the United States of America’s status as the foremost nation in health care innovation.”


I’ve been a distracted mess lately, with all this PT stuff as well as a week of administrative malarkey, but I did notice a a provocative comment that I feel compelled to respond to.
Upon accepting the risk of dispensing an unpopular remark: One day, we shall have to set nature in order using genetical engineering.
No creature should devour any other.
Wow. That makes no sense. We humans are obligate heterotrophs — we must obtain certain vital molecules by consuming other organisms. For example, we cannot synthesize valine, isoleucine, leucine, methionine, phenylalanine, tryptophan, threonine, histidine, or lysine, so we have to consume other organisms that contain those substances, or we die. I guess we can define “creature” to escape the problem, which is the vegetarian solution. We don’t eat meat from animals by making the decision that plants don’t count. It’s very convenient to say that killing carrots or yeast or lettuce is ethically OK, but if you think about it all deeply, even a carrot is a product of processes that kill insects with pesticides. Do insects count? What about protists? The lines are all arbitrary and we each draw our own lines.
Is our solution to genetically modify humans so they can synthesize every molecule we need? Or are we going to build factories to create all these essential substances as supplements?
But deeper still, the planetary biome is built on dependencies contingent on death and consumption, in every food web that exists. For example, sea otters eat sea urchins; sea urchins eat kelp; when sea otters are eliminated, the kelp forests die. How do we genetically engineer “devouring” out of the system without necessarily deleting entire ecologies? The only way any of this can happen is by magic.
Nature is already in order, reordering it to your preferences is silly.

Liang Cheng is an oncologist a Brown University. I’d never heard of him before, but I am told that he is incredibly famous by Liang Cheng, as he announced himself on LinkedIn.
I am deeply humbled and grateful to learn that my H-index has now reached 140. I was also honored to see that I am currently ranked among the two most-cited researchers worldwide in the fields of Urologic Oncology and Urology on Google Scholar.
In addition, my i10-index has reached 1060; that is, one thousand and sixty publications each cited at least ten times. I was told that this may represent a world record – what an extraordinary honor!
Nonetheless, these numbers are far less important than the people, mentorship, friendships, and collaborations behind them. This milestone is truly a triumph of team science. I owe immense gratitude to my mentors, colleagues, collaborators, residents, fellows, medical students, and friends who have inspired and supported me throughout this journey over the past two decades.
Academic medicine is never an individual accomplishment. It is ultimately about advancing science and medicine, educating future generations, and improving patient care. If our work has contributed even in a small way toward those goals, then I feel extraordinarily fortunate and grateful.
Thank you for being part of this journey. The best is yet to come!
I hate to be the one to tear him down, but no one cares about your H-index and i10-index except, maybe and importantly, administrators and fellow H-index chasers. Anyone else remember that scene in American Psycho where Patrick Bateman and several of his cronies are comparing business cards, noting the quality of the stock and the embossing and the inks? Yeah, that’s what it’s like seeing someone brag about their indexes. Don’t care.
It’s also because those numbers are thoroughly gamed. I looked him up on PubMed, and it’s true, his name is on a lot of papers: papers that have 10 or 20 or more authors, and there he is, somewhere in the middle of a sea of names, rarely first or last. He really does owe a lot to his “mentors, colleagues, collaborators, residents, fellows, medical students, and friends” who have been tacking his name unto their papers! And further, his publication rate, that is, the rate at which his name gets plugged in to a long list, is approximately a paper every two days, which is insane. This is authorship by rubber stamp.
I think it is valid that many research endeavors nowadays require a large team, and he may have been an indispensable member of such a team, but then to use that cooperation to brag that he is #1 or #2 in his field is unseemly. It’s also dangerous, Dr Tall Poppy. He was spamming his ‘accomplishment’ on every social media site he could find, and on Xitter, Michael Eisen noticed.

The author’s Google scholar profile falsely lists multiple papers that he didn’t author, and therefore the citation count and h-index are inaccurate.
Whooops.
I do enjoy seeing a braggart taken down a peg, but Liang Cheng is a symptom of a greater problem: we’re drowning in artificial metrics, amplified by AI slop.
Over the last few decades, science has undergone a “citation revolution.” Scientific life used to be structured by personal reputation and mutual acquaintance; now it is defined by quantitative assessments derived from citations.
And this reward system has warped scientific life in dramatic ways. It has resulted in the obvious and widespread gaming of citation metrics; but, more insidiously, it has pushed scientists toward risk-averse, incremental, and above all unambitious research. The logic of institutional science has become increasingly divorced from actual knowledge and discovery. In a system governed by these perverse incentives, the inevitable endpoint is simply AI-generated slop at scale.
Now, with AI, we’ve built a remarkable new technology that opens up dramatic new horizons for scientific work. But we’re deploying that technology within an institutional structure that incentivizes, above all else, the maximization of metrics that don’t have much to do with real science. The underlying problem is not with AI, but with the institutions and incentives that define modern science.
That is an excellent article, everyone should read it. It actually ends on a promising note, regarding AI as a tool that could break us out of the dead-end, grasping competition for a magic ranking number, as exemplified by the case of Liang Cheng.
The citation index was designed in the 1950s and ‘60s as a solution to the information crisis engulfing scientific life. It ended up becoming much more than that: a regime that reshaped what science was, how it was rewarded, and what kind of science got done. Now that regime is collapsing under the weight of its own contradictions. I think it’s a fantastic opportunity to build something better.


A scientist has been denied access to his lab…because he supported his Chinese students.
A faculty member at Indiana University (IU), who has sharply criticized the government’s recent prosecution of several Chinese scientists accused of smuggling biological materials into the United States, has been locked out of his laboratory by the school in response to a request by one of his federal funders.
IU plant microbiologist Roger Innes says the move Thursday evening is the latest instance of retaliation for a letter he wrote last fall on behalf of Yunqing Jian, a plant scientist postdoc at the University of Michigan who had pled guilty to smuggling biological material and making false statements. The letter to Jian’s attorney, intended to be used at her sentencing, argued that what the Chinese postdoc had transported was not dangerous, but she was still ultimately deported. Her conviction triggered an investigation of Youhuang Xiang, a Chinese postdoc in Innes’ lab, that led to Xiang also pleading guilty last month to smuggling loops of DNA known as plasmids. He was also deported.
Oh no, never criticize the US government.
This sounds like overreach by government agents trying desperately to find excuses to deport Chinese scientists, and taking out an American scientist as collateral damage. If I had foreign students, I would defend them without question, but apparently that will get you shut down in America.
I would love to know the rationale for excluding the PI from the lab, even if his post-docs were guilty of importing nefarious plants. Do they suspect him of plotting to attack the US from his lab in Indiana with evil weeds from China?
Interesting touch: the chair of Innes department is Armin Moczek, an eco-evo-devo guy I know of. He’s going to be watering Innes’ plants while he’s locked out.

It’s the perfect storm, and I have myself to blame. I gave the fourth exam of the semester yesterday, and many of the students bombed. This happens every year, and it’s because the fourth exam is the hardest of them all: everything accumulates in this course, and it’s the combination of lots of math and understanding the topology of weird chromosomes entangled and swapping bits around that breaks their brains. Couple the fact that these are generally smart, high-achieving students with the shock of getting a 40% on an exam, and it’s going to get ugly. I’m going over the answers with them this afternoon.
Also, we have a major lab report due next week, and an in-class presentation. It is a high stress situation.
Students are already emailing me with requests for a point here and there on back assignments and exams. I’m going to spend my weekend with spreadsheets recalculating all their old assignments and putting together a final assessment, which will inspire a further frenzy of grade-grubbing.
If I drank anymore, this would be the weekend I’d finish off a bottle of good whisky, but I don’t drink at all anymore. I’d rather talk about genetics than grades, but the system will not allow that.


I lost all respect for Matt Ridley years ago, when I wrote:
Matt Ridley is definitely a smart guy, and he also writes well. I enjoyed some of his earlier books, like The Red Queen and Genome, but I became less appreciative as he became more openly libertarian, and espoused a Whiggish view of the world that was only a rationalization for why he was so wealthy and privileged (he’s kind of the British version of Pinker, only worse). He’s the 5th Viscount Ridley, don’t you know, he is to the manor born (Blagdon Hall, Northumberland, specifically), he’s a member of the House of Lords, he endorsed Brexit, he owns coal mines, he used to own a bank, but he ran it into the ground and it was taken away from him and nationalized. On climate change, he’s argued that global warming is going to be a net benefit, increasing rainfall and the growing season, and that human ingenuity will overcome any minor disruptions. He even coauthored a book with Anthony Watts and Bjorn Lomborg and a host of the usual denialist suspects, Climate Change: The Facts 2017, which ought to alarm anyone who wants to think he’s just being objective. I guess that comes of owning coal mines and being an enthusiastic endorser of fracking — when your prosperity is a product of spewing as much fossil carbon into the atmosphere as you can, your very smart brain will work very hard to find excuses.
At this point, he is an irredeemable kook — but a well-connected and wealthy one, who gets invited to all kinds of events hosted by the corrupt, criminal kooks in charge of the US government. He was recently invited to address the NIH on the “lab leak” hypothesis, the discredited conspiracy theory that China intentionally engineered the COVID virus to wreak havoc on the West, but accidentally released into their home territory of Wuhan. It’s absurd. No one who knows anything about virology or molecular genetics thinks it is at all plausible, or credits it as valid in the face of all the evidence that it originated naturally from wild populations, or zoonosis.
Nevertheless, Ridly got invited to present his innuendo, lack of evidence, and leaps of illogic at a major meeting of the formerly prestigious NIH.

Must Trump get his face put front and center of everything?
Don’t trust me that the “lab leak” is a garbage hypothesis? Larry Moran and Lindsay Beyerstein both shredded these “lab leak” claims years ago, but conspiracy theories seem to be invulnerable to little things like evidence and reason.
Add another critic to the long list of knowledgeable scientists who find Ridley risible: Angela Rasmussen. She gives five really strong reasons why the “lab leak” nonsense is wrong, and also seems to have even less respect for Ridley and Bhattacharya than I do.
I don’t know Ridley’s motivation besides being a pompous literal coal baron who led the UK into the 2007 financial crisis with the first run on a British bank in 130 years, and who expects to be taken seriously despite the fact that he seems like a foppish secondary antagonist in a forgettable Dr. Who episode. Unfortunately, Ridley is taken seriously by his fellow pompous, insubstantial windbags, and even more unfortunately, they are the windbags in charge who invited him.
It is no mystery why Podcast Jay [Bhattacharya] rolled out the red carpet for the distinguished Viscount despite his lack of qualifications or relevant expertise on the topic. He knows that Matt Ridley is writing fiction. He shows up to launder conspiracist lies through an indignant upper class British accent in service of Bhattacharya’s ultimate goal: to declare all NIH research reckless, dangerous, and absolutely the worst thing we could possibly do. Better redirect the money for these irresponsible Alzheimer’s, HIV, and diabetes treatments to the White House. President Donald Trump and Office of Management and Budget Director Russell Vought will know what to do with all those funds.
I don’t see enough news and criticisms of Bhattacharya — he’s in charge of the NIH, he’s an idiot, he’s busy dismantling the American scientific institutions, but mostly what gets into the press are the disastrous decisions of the bigger fools in the cabinet. Make no mistake, though, he’s one of the nastier parasites gnawing at the foundations of our science establishment.
And now he invited Ridley to speak. The Ridley who makes these kinds of ludicrous accusations:
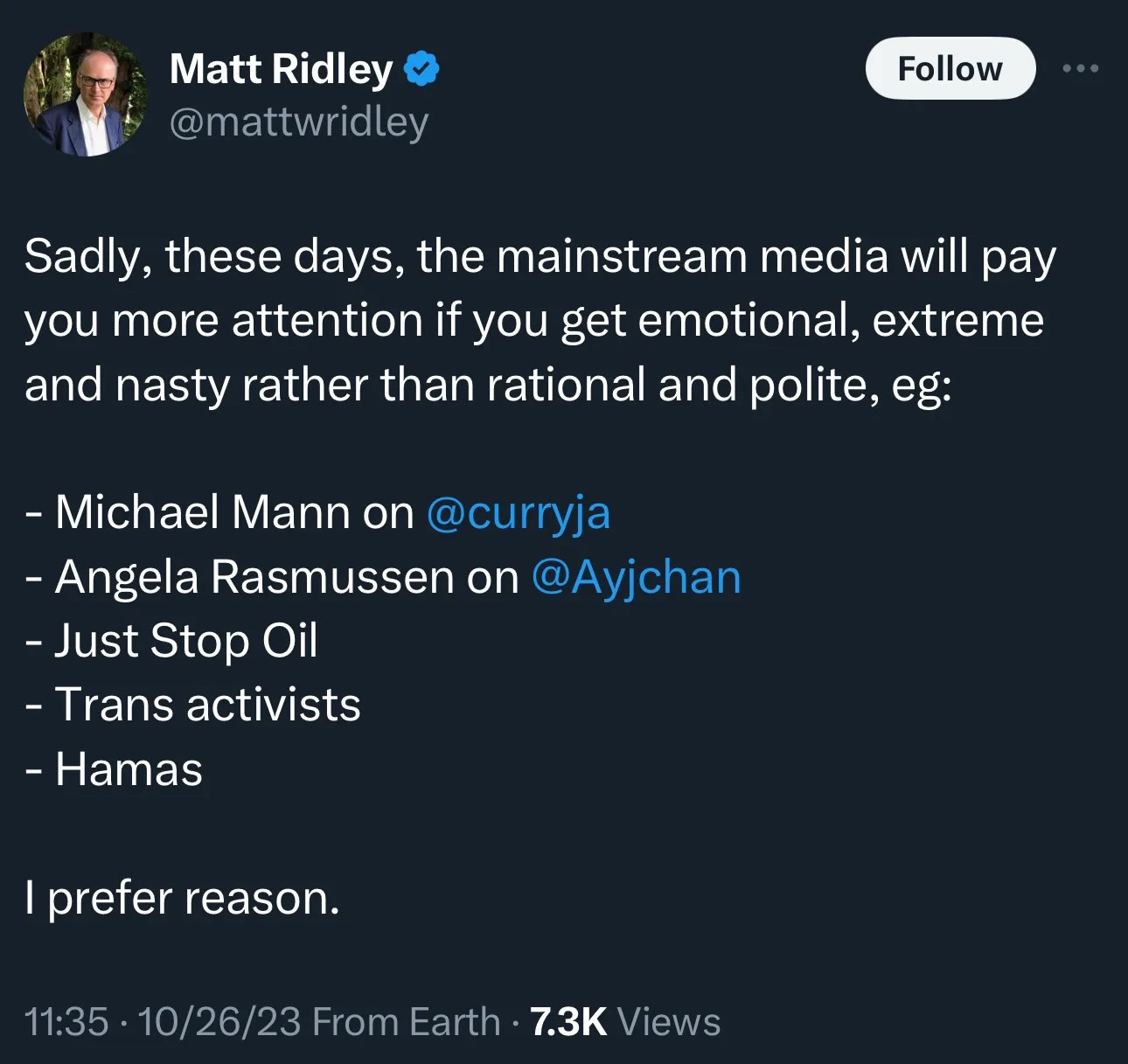
You can tell what kind of man he is by the people he lumps together as enemies with Hamas: a couple of credible, qualified scientists and trans people. His right-wing politics are showing.
