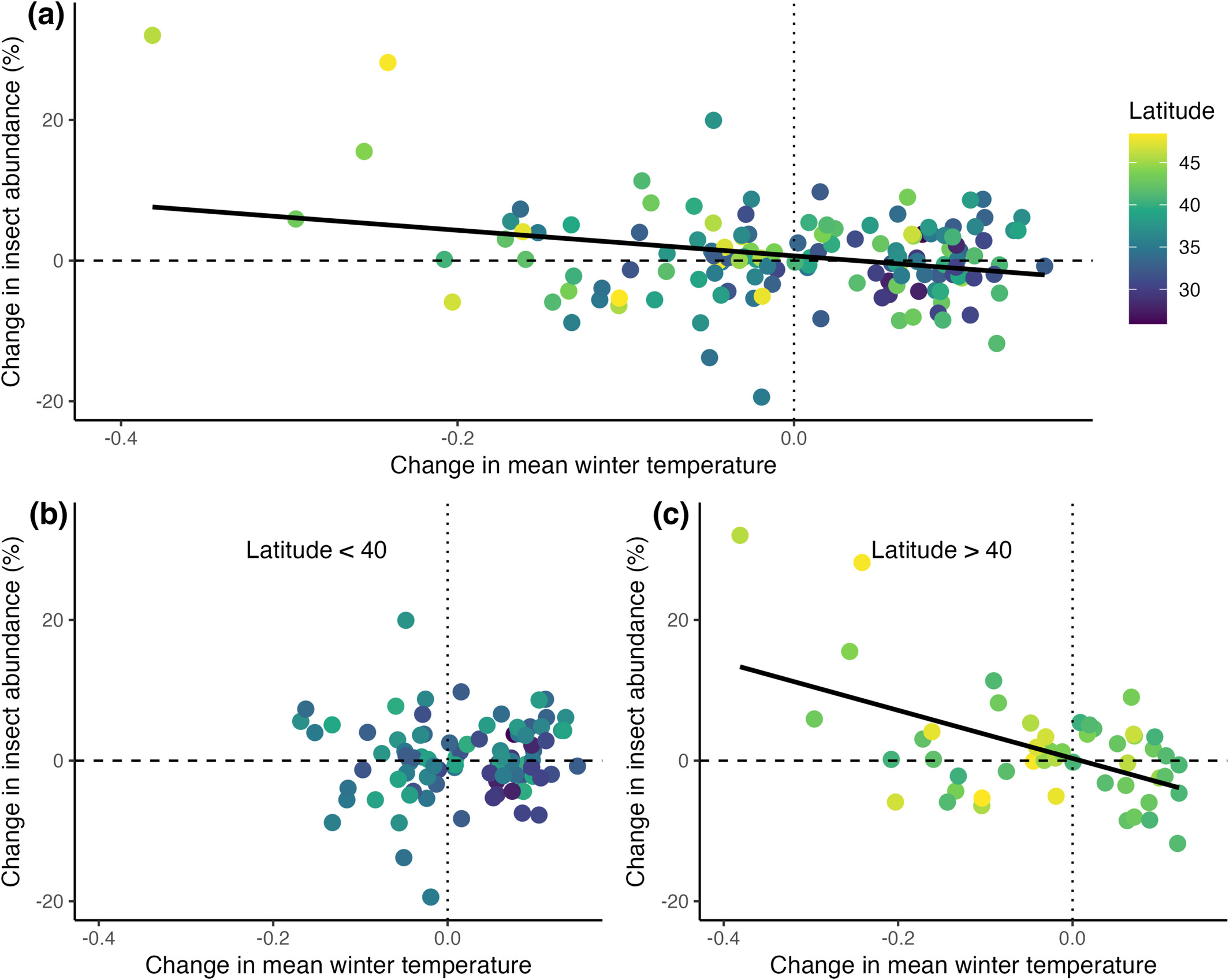
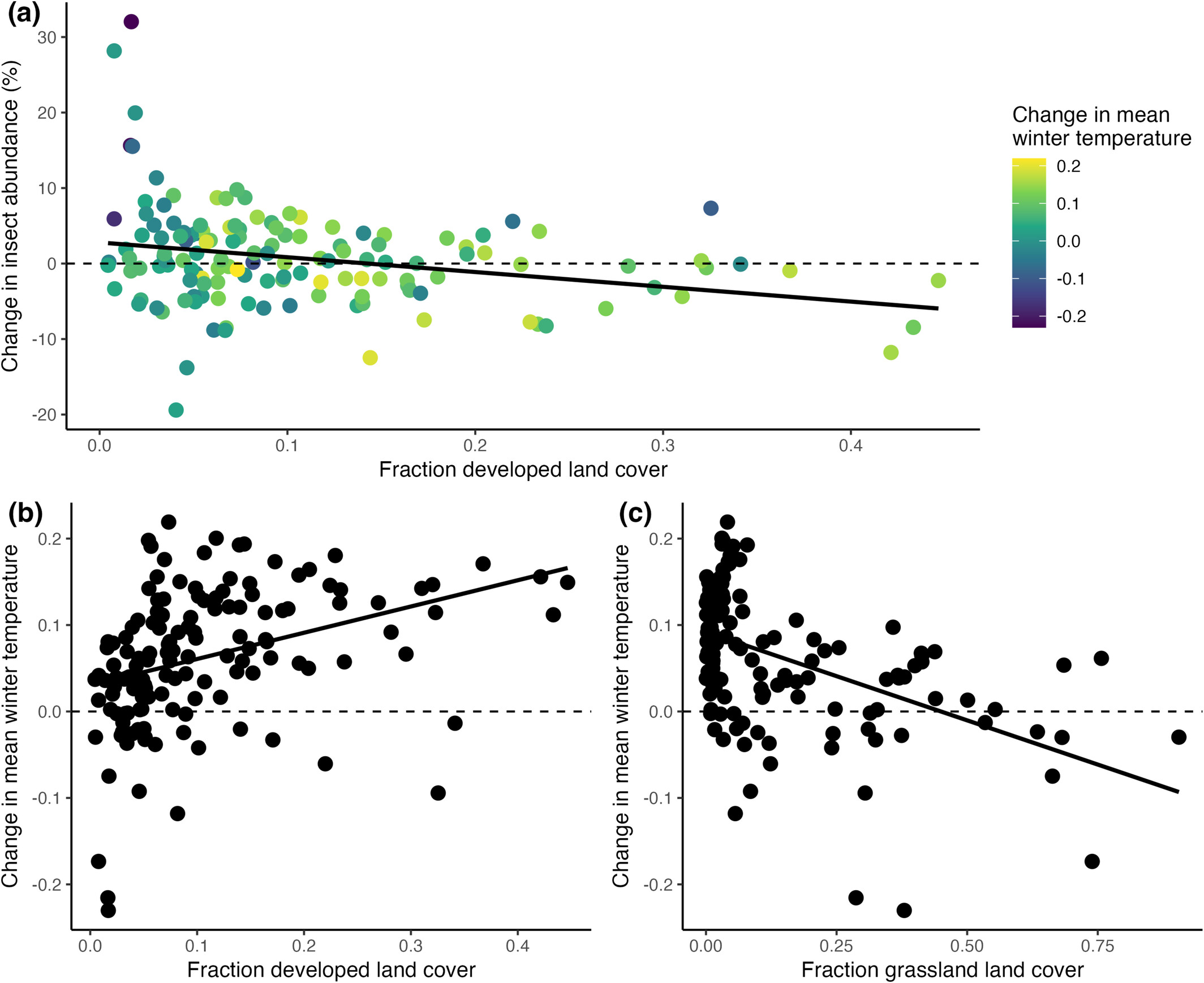
I’m a terrible YouTuber — uncharismatic, dull, lacking in visual skills, and incapable of maintaining a consistent schedule — but heck, I’ll try again. Tomorrow (Saturday, 20 December), I’ll go live around 3pm Central time. I’m open to talking about just about anything, but will center the discussion on this paper:
Christopher J. Kay, Anja Spang, Gergely J. Szöllősi, Davide Pisani, Tom A. Williams & Philip C. J. Donoghue (2025) Dated gene duplications elucidate the evolutionary assembly of eukaryotes. Nature, 3 December 2025, DOI: 10.1038/s41586-025-09808-z.
If you don’t want to read an 11 page technical paper, just contemplate this figure:
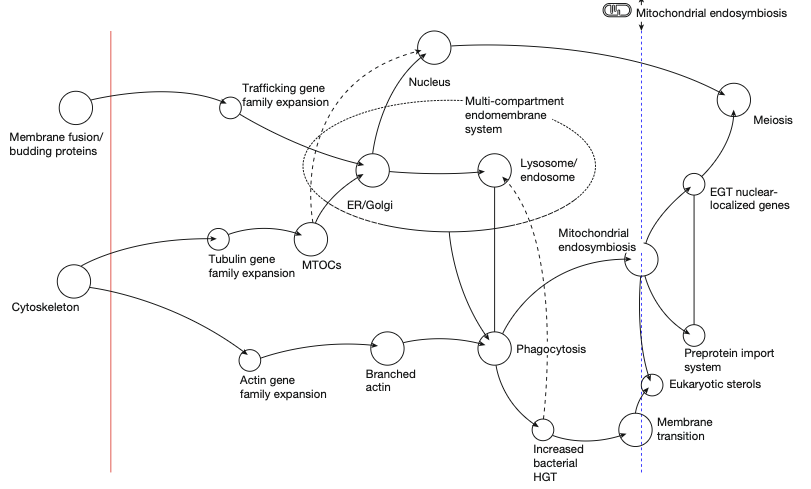
Or you can just tune in and maybe I’ll explain it.