In this post I will be looking at the rest of the proteins in stage 1 of purine biosynthesis, the related PurL and PurM. To this point an ammonia dispenser in PurF provided an ammonia that replaced the diphosphate on ribose in PRPP, an ATPgrasp ATP phosphate dispenser used a phosphate to add a glycine, and then a formate was added to the nitrogen on glycine with PurT/PurU, or PurN. This leaves us with FGAR, formylglycinamideribonucleotide.
Now I add steps 4 and 5. Swapping the glycine carbonyl with an imine, and closing the first purine ring.
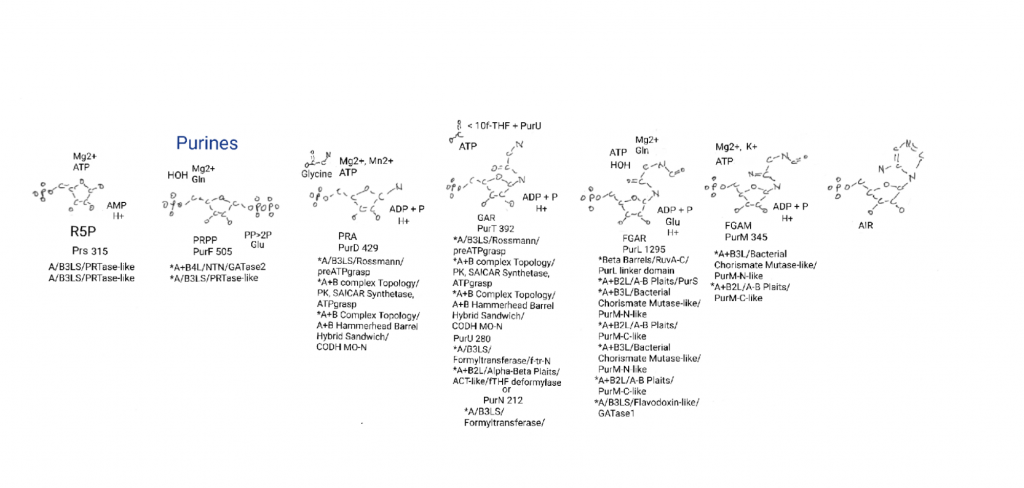
Step 4: PurL and swapping the glycine carbonyl with an imine via phosphate.
PurL is a large protein, the largest on the drawing. To me large and usually complex means old. This protein uses a phosphate from ATP to swap the glycine carbonyl oxygen (C=O) for an imine (C=N). The ammonia again comes from glutamine. Even though it is probably old parts of it can be deleted for fun if I assume ammonia is in the vent.
PurL has 5 individual domains, 2 present in duplicate. I consider some parts more important than others. In the vent the entire glutamine ammonia dispenser might be unnecessary.
- An Alpha Arrays/RuvA-C-like domain (needs corrected from Beta Barrels on the drawing), the PurL linker domain.
- An A+B2L/Alpha-Beta Plaits domain, the PurS-like domain.
- An A+B3L/Bacillus chorismate mutase-like/PurM-N-like domain (present in duplicate).
- An A+B2L/Alpha-Beta Plaits/PurM-C-like domain (present in duplicate).
- An A/B3LS/Flavodoxin-like/Glutamate amidotransferase class 1 domain (GATase1).
RuvA-C-like domain
Starting with the Alpha Arrays/RuvA-C-like domain. This is a linker domain that acts in long range communication between parts of the protein. It’s all by itself in its homology group so that’s that.
PurS-like domain
This second domain in PurL is an Alpha + Beta 2 Layers domain called “PurS-like” because it is sometimes a separate protein, but always required for PurL function so in that case they call it “PurS”. It’s function isn’t well known but it isn’t the part that makes ammonia or holds FGAR, maybe it just holds the other 2 domains together. This is also in its own homology group all alone.
PurM-N-like domain
This domain is in the “bacillus chorismate mutase-like” X group, which may be related to the N-terminal part of PurM/parts of PurL. And maybe not. There is an interesting set of things in here and I consider this part and the following PurM-C-like domain to be the most important.
First is HypE which is involved in maturation of hydrogen utilization proteins with complex iron-nickel centers, hydrogenases. They can turn H2 into 2 protons and 2 electrons and back again. When it comes to the concept of energy, cells need to have and control a pool of protons and electrons, using things like niacin and riboflavin.
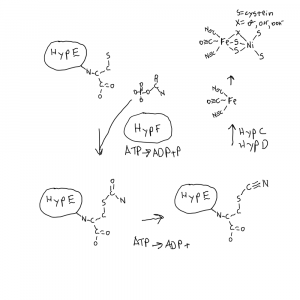
HypE binds a carboxamido (N-C=O) on it’s C-terminal cysteine, that came from carbamoyl phosphate (as in pyrimidine biosynthesis), and delivered by HypF. ATP is then used to dehydrate it to a cyanide (nitrogen triple bonded to carbon). This cyanide and a second is then bound to an iron atom (On HypC-HypD) used to make the finished iron center that is combined with a nickel atom.
Structural Insight into [NiFe] Hydrogenase Maturation by Transient Complexes between Hyp Proteins Miki 2020
Selenide, water dikinase, also known as selenophosphate synthetase, SelD. Synthases and synthetases both ligate (connect) things together but synthetases need a source of energy like ATP.
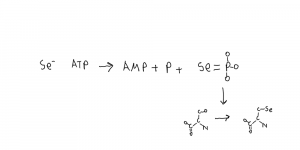
This combines selenium and phosphate to make selenophosphate which is the form used by cells. Selenophosphate is then mostly used as the 21st amino acid in a system that replaces serine with sersec (Sec, U) on the tRNA, or an RNA modification. The serine translation system is used and individual sec replacements are made during translation, hence the amino acid sersec. This is in bacteria, archaea and eukarya.
The selenophosphate synthetase family: A review Manta 2022
Finally thiamine monophosphate kinase (ThiL, on the big drawing) makes mature thiamine diphosphate.
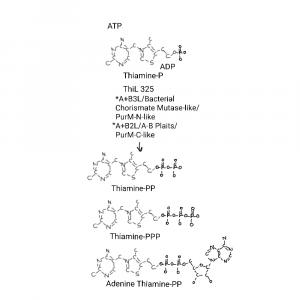
I type mature because as far as I have found thiamine and thiamine monophosphate do nothing. It is thiamine diphosphate that is mostly used by cells, with some thiamine triphosphate and adenosine thiamine diphosphate. Those last 2 are somehow associated with glucose availability and amino acid starvation, and the opposite, amino acids and no glucose, respectively.
Update on Thiamine Triphosphorylated Derivatives and Metabolizing Enzymatic Complexes Bettendorff 2021
PurM-C-like domain.
And just like that we’re done with this domain because again we have PurL, PurM, HypE, selenophosphate synthetase, and ThiL. The only difference is this part of the protein is an Alpha+Beta 2 Layers and then in the Alpha-Beta Plaits X group, within which there is no guarantee of relationships outside of the general alpha helix and beta sheet organization.
Class 1 glutamine amidotransferase domain.
This is the glutamine ammonia dispenser for this protein. GATase class 2 we saw with PurF, a GATase2. The larger family this is a part of, the X:Flavodoxin-like and H: GATase1-like A+B3LS domains, appears many times on the big drawing. There is a lot to point out here. 20 related topology bins are siblings with the GATase1-like bin. And that topology bin has a lot besides the GATase family.
First the GATase1-like topology group itself has a lot of things already on the drawing. The current PurL, GuaA, CarA, PyrG, AroE, TrpG, PabA, FolD HisH, MetA and Pdx2. There are more but they are in different topology bins. This topology group also has protease 1, peptidase S51, peptidase C26, peptidase S66, catalase 1 (breaks down peroxide), isocyanide dehydratase, and beta-galactosidase.
The same topology bin as AroE and FolD (amino acid dehydrogenase-like-N) also contains Glu/Phe/Trp/Leu/Val (amino acids) dehydratases, malate oxioreductase (makes pyruvate from a TCA cycle intermediate), and at least 2 pterin (folate relative) proteins, methylene tetrahydromethanopterin dehydrogenase and tetrahydrofolate dehydrogenase/cyclohydrolase.
Pterin related, in its own topology bin is F420 dependent methylene tetrahydromethanopterin dehydrogenase.
In separate topology bins and also on the big drawing we have PurE, RibH, AroB (in a bin with glycerol dehydrogenase), and AroQ.
Things that grab things and move them is a theme for the Periplasmic Binding Proteins-like 1 and Chelatase- like bins. In bacteria with double membranes the space between them is called the periplasm. Periplasmic binding proteins bind things like amino acids (leucine here) and sugars (glucose, galactose, arabinose here). Chelatases bind and insert tetrapyrroles like heme and chlorophyll, which in turn often bind, “chelate” metal ions. This bin also contains iron-molybdenum binding proteins, zinc transporters, cobalt chelators, vitamin B12 transporters, a iron chelators.
Relatedly in its own topology bin is uroporphyrinogen 3 synthase, or HemD. This protein turns a linear chain of 5-membered rings into a circular ring heme/chlorophyll precursor, uroporphyrinogen 3. These often feature a chelated metal ion in mature form.
A last grabber/holder is the ATC-like topology bin, where the “ATC domain” contains a bunch of cysteines in the sequence which likely hold iron-sulfur centers, and is related to aspartate and glutamate racemases.
In a topology bin by themselves is phosphofructokinase and NAD (niacin) kinase. A glycolysis protein and the protein that makes NADP from NAD, and thus switches niacin to the anabolic form from the catabolic form.
The CheY-like bin is named after a chemotaxis protein, and contains pili related proteins, HydB (a NiFe hydrogenase), sensor kinases (RcsC), a vitamin B12 binding domain which is in MetH (big drawing), and ornathine decarboxylase.
In a topology bin by themselves there is fucose and arabinose isomerase. Another separate topology bin contains YfiR which makes cyclic di-GMP. Yet another topology bin contains IlvD (isoleucine, valine, and coenzyme-A biosynthesis) and 6-phosphogluconate dehydratase.
And finally the interesting FabD/lysophospholipase-like topology bin contains acyl carrier (FabD) and transfer proteins. It also has parts of fatty acid synthase. Binding and carrying coenzyme-A is a big theme. Lysophospholipases separate fatty acids from the glycerol backbone.
Step 5: PurM
PurM closes the ring via a phosphate from ATP, and the same basic reaction mechanism as PurL.
Thanks to PurL, I’m done with PurM since it’s the same 2 fragments shared with HypE, selenophosphate synthetase, and ThiL.
That’s it for stage 1 of purine biosynthesis, forming the first ring.
Fascinating read! Your explanation of PurL and its role in purine biosynthesis is clear and insightful. Excited to see what you cover in Part 3!
dude theft auto
The best part about Wacky Flip is that it never gets old. The randomness keeps it exciting every time you play. You’ll always want to beat your best flip.
Paper io transforms the battlefield into a contest of precision and strategy. Players expand their territory by drawing lines that turn into land once safely completed. While doing so, they must watch for rivals ready to strike their exposed trails. Paper io encourages strategic thinking, map awareness, and calculated aggression. Its clean visuals and smooth controls allow players to focus entirely on movement and decision-making.
But while single-word guessing games have dominated the spotlight, another type of brain teaser has quietly been winning people over: Phrazle, a game built around complete phrases instead of just one word.
Dense but fascinating, nice breakdown of how PurL and PurM tie metabolism together, especially the recurring themes of ammonia transfer and ATP-driven chemistry
The breakdown of PurL into distinct domains is helpful, but I wish there was more detail on how each domain contributes to the overall mechanism. Are there structural studies that show domain interactions during catalysis?
The racing game Kart Bros. is a strategic racing game that helps players refine their skills to analyse the circuit and alter their strategies after each acceleration. Not only does each race involve racing speed, but it also necessitates the ability to select appropriate steering angles and devise strategies for overcoming obstacles.