I’m having big motivation issues and it’s making it harder to mentally assemble my next post so I thought I would do my reading list and share a couple of the things I want to work into the post. Hopefully this will jog things around. The stir crazy always comes back, it’s taking its time this time.
Understanding Emotions: Origins and Roles of the Amygdala.
In the last 2 years I did a binge on the amygdala in general to get a good feel for that part of the brain. It works as a “cognitive insertion point” which I think of as a place where you can draw on individual experiences to discuss a part of the brain. The amygdala receives and processes objects, and inserts direction and intensity of feelings into consciousness as well as a bunch of instructions for movement. It’s a “system 1” level structure as it requires no effort, you just feel it. A “system 2” structure is involved in effort in consciousness.
Novel Perspectives on the Development of the Amygdala in Rodents.
I finally decided to get more developmental concepts internalized. A flesh tube becomes brain parts and some of those parts migrate to make new brain parts. The amygdala is an “interface structure” and made from lots of meat that wandered in from elsewhere and sits next to things that also wandered in from elsewhere.
Development and function of the medial amygdala.
I paid particular attention to the central amygdala when I did my binge (CE or CeA). Over 90 links to articles on just that region. It’s the output subnuclei of the amygdala. The medial nucleus (ME or MeA) outputs to the CE and seems to be a junction between the senses and behavior. Stimulation of the CE produces strong salience (attention) to the point that rats will chew at an electrode that shocks them and so this nuclei features in addiction research. The ME when stimulated produces many kinds of behavior. (And in my last post I pointed out the posterior cortical nuclei seem to create feelings).
Organization of neural circuits underlying social behavior: a consideration of the medial amygdala.
Same thing but with a social behavior focus. Oxytocin discussion and how to tease apart the sensory from behavior parts of this nucleus. Since body maps are involved in movement maybe there’s a sensory body component that works with cuss words?
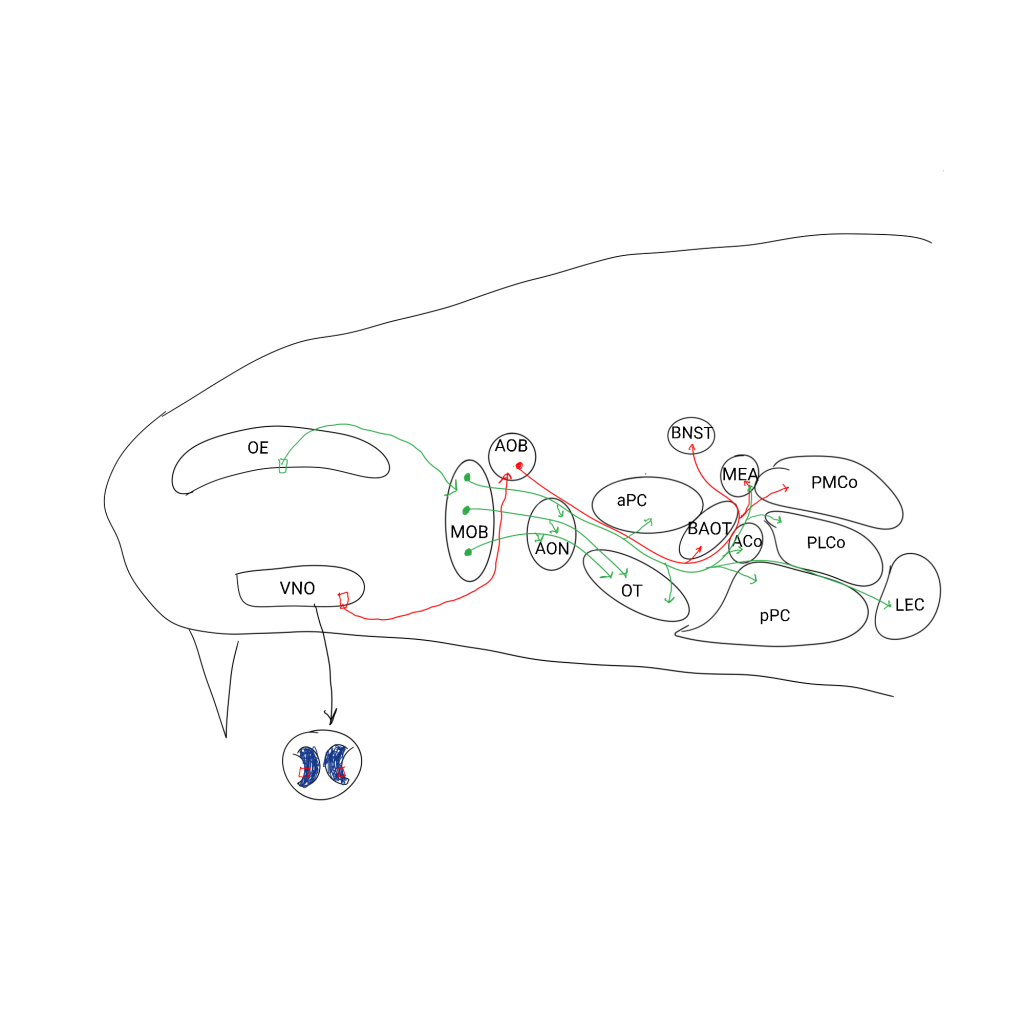
TRICK or TRP? What Trpc2−/− mice tell us about vomeronasal organ mediated innate behaviors.
Related to my last post. TrpC2 is a gene used in the vomeronasal organ and makes the accessory olfactory bulb severely reduced when deleted in mice. It also eliminates social aggression and effectively makes them pansexual. Female mice even scent mark like males in addition to mounting both sexes and thrusting away. The human TrpC2 is a pseudogene.
I want to post about these mice but I want to read more about a surgical counterpart, VNX mice with surgically removed vomeronasal organs.

This is an outline of how the stream of consciousness, feelings, and emotions seems to relate as a continuous looping process. And how attention can be moved around in that stream. We experience the world as continuously cycling and updating networks of neurons.
Emotions seems to come in approach-hold-withdraw kinds (or attraction/avoidance…), and fight-freeze-flight for an intensity spectrum. There is an emotional motor system that seems to govern approach and withdraw based on feeling. This feeds into what goes to the motor system proper.
What I want to do is remake this with the anatomy.
The next part came from some notes I took after reading about the structure of the autonomic nervous system. I’m trying to define and categorize all of the internal senses and feelings in preparation for finding them in the anatomy. If I missed any let me know! This should be things we all feel. It gets a bit repetitive as I’m looking at different ways of looking at the same thing, or just listing things to get them all.
Sense of Parts
Organs, muscles, bones, skin…
Sense of State
1) Sense of place in space: Proprioception
Special case: non-localized/soft/crude touch.
2) Sense of self-directed movement
Sense of Metabolic State
3) Sense of Nutritional State (hunger, specific hunger, thirst, starving…)
4) Sense of Digestive State (feeding, digesting, nausea, waste elimination)
5) Sense of Physical Integrity (Injury)
6) Sense of Chemical Integrity
Physiological (immune, O2, CO2, pH, pressure, temperature, hormonal…)
Arousal
Emotional state relative to (self, other beings, environment), feelings about.
External Senses
Smell
Vision
Hearing
Taste
Touch
Rotation
Acceleration, movement, and balance
Internal Senses
Internal parts: Introceception
Visceroception: sense of organs and muscles
Position and movement of parts: proprioception
Internal state: autonomic sense
Hunger
Specific Hunger
Thirst
Digestive distress
Waste elimination
Injury
Alertness/arousal and changes to (social hunger)
Heart rate, respiration,
O2, CO2, pH
Temperature
Immune system status
Emotional state relative to X