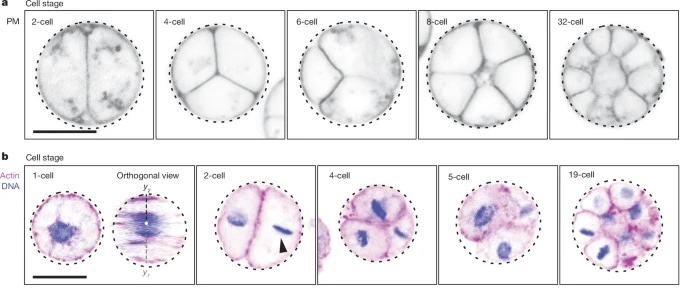
I teach (in the distant future) a first year class in genetics and evolution. I’m going to have to use this comic when I discuss the inheritance of acquired characteristics — I think the bit about helicopter moms might resonate.

But not now! No teaching until January 2026! I’ll just file this away for now.