Seahorses are weird animals. They depart from the typical streamlined torpedo shape of your average fish to construct this unusual twisted shape with dermal armor, toothless jaws, and a dependence on fins for propulsion — they’re just weirdos all around. How did they get to be this way?
One suggestion is that it is an extreme example of paedomorphosis, as presented in this paper: An embryonic arrest shapes the Syngnathid body plan: Insights from Seahorses, Pipefishes, and their Relatives.
The Syngnathidae (seahorses, seadragons, pipefish, pipehorses) exhibit a remarkable, enigmatic body plan, challenging conventional explanations for their fused jaws, toothlessness, cartilaginous skeleton, fin loss, male pregnancy, and their distinctive morphology, which includes the acute head-trunk angle of seahorses and the family’s unique curling, often prehensile, tail. We propose a unifying, parsimonious hypothesis, termed “pharyngulation,” that the entire lineage originated from a profound paedomorphic arrest (retention of juvenile traits) during a specific embryonic pharyngula stage. This arrest, likely driven by ancestral Hox gene cluster disintegration, fundamentally halted morphological progression in a common teleost ancestor. This single event explains their entire suite of primary characteristics–including universal low body mass and volume and unique A-P locomotion. It also establishes a framework to differentiate these foundational family-defining traits from ancestral features shared with the broader Syngnathiformes order (such as the elongated snout, as exemplified by Trumpetfish) and from later adaptive refinements, such as the leaf-like appendages in seadragons. Our “pharyngulation” hypothesis offers a novel, testable model for macroevolutionary innovation, demonstrating how a singular, profound alteration to a conserved developmental program can rapidly forge a new, viable body plan. This concept, synthesizing evidence from genomics, the fossil record, and developmental biology, is of broad interest to evolutionary biologists and developmental biologists alike.
Unfortunately, this paper only presents a hypothesis — no methods, no experiments, no substantial comparative data. I’ll forgive that since it does introduce the term “pharyngulation” into the scientific literature.
I was provoked to dig a little deeper, and found this paper: A comparative analysis of the ontogeny of syngnathids (pipefishes and seahorses) reveals how heterochrony contributed to their diversification. It supports some of the ideas of the first paper — heterochrony is right there in the title — and also includes some beautiful photos of syngnathid embryos.
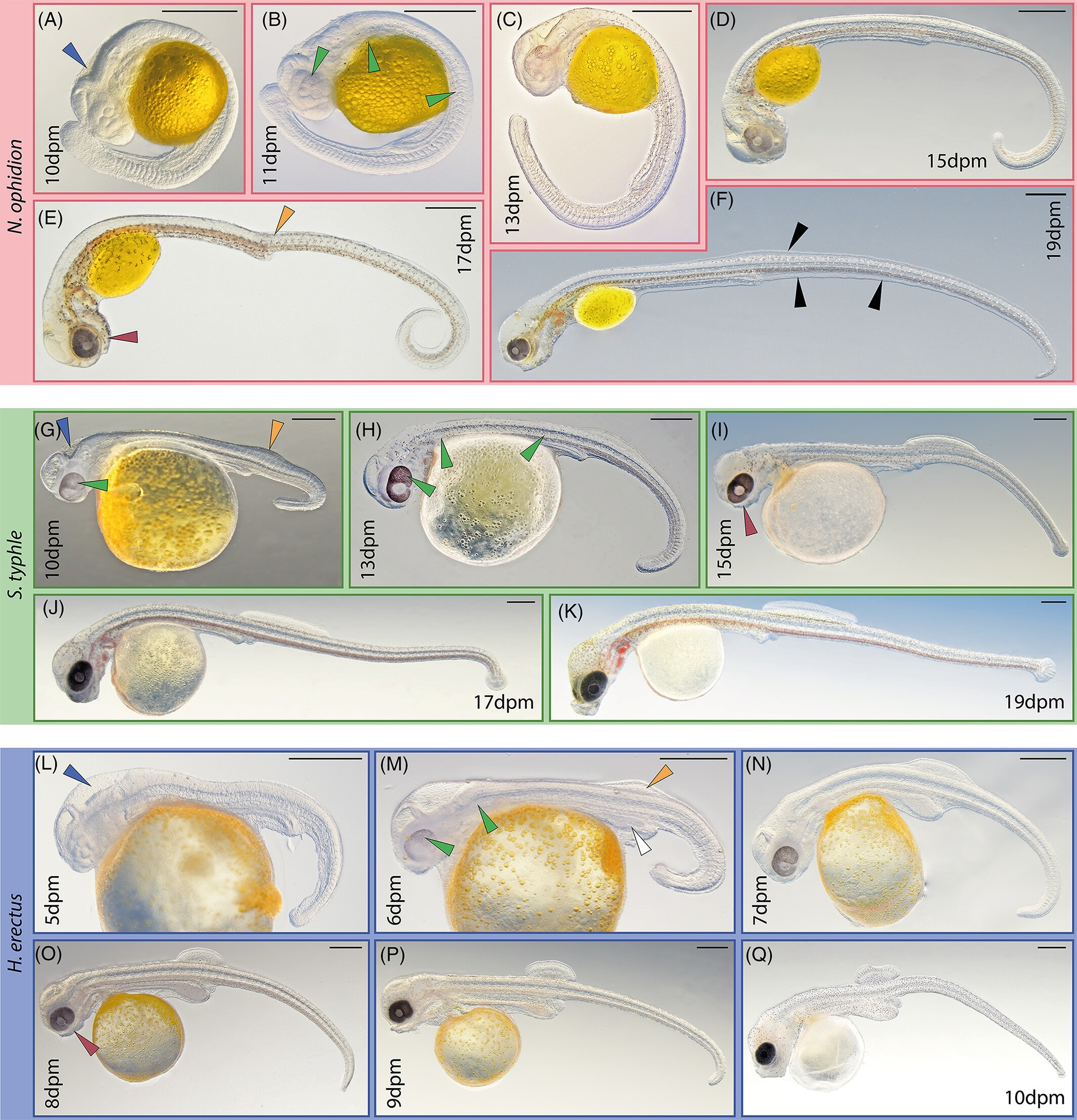
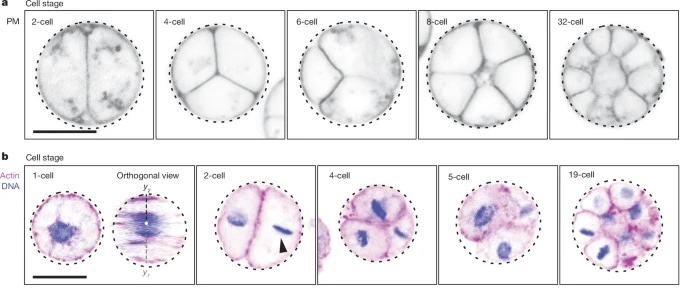
Segmentation and early organogenesis development in examined syngnathids. Nerophis ophidion (A-F), Syngnathus typhle (G-K), and Hippocampus erectus (L-Q), respectively. In this period, species-specific characteristics develop more clearly. Arrowheads: blue = hind brain vesicle, green = pigmentation, rufous = mandibular arch, orange = dorsal fin condensations, white = hypertrophic hindgut, black = fin fold. Scale is 500 μm; dpm = days post mating
That’s a stage close to what we’d call the pharyngula stage (which doesn’t have a single discrete marker), and they look familiar — they look like longer, skinnier, more slowly developing zebrafish embryos, where the 19 day syngnathid looks like a 19 hour zebrafish. We have to wait a week or more to see an embryo that is comparable to, but very different from, a 24-48 hour old zebrafish embryo.
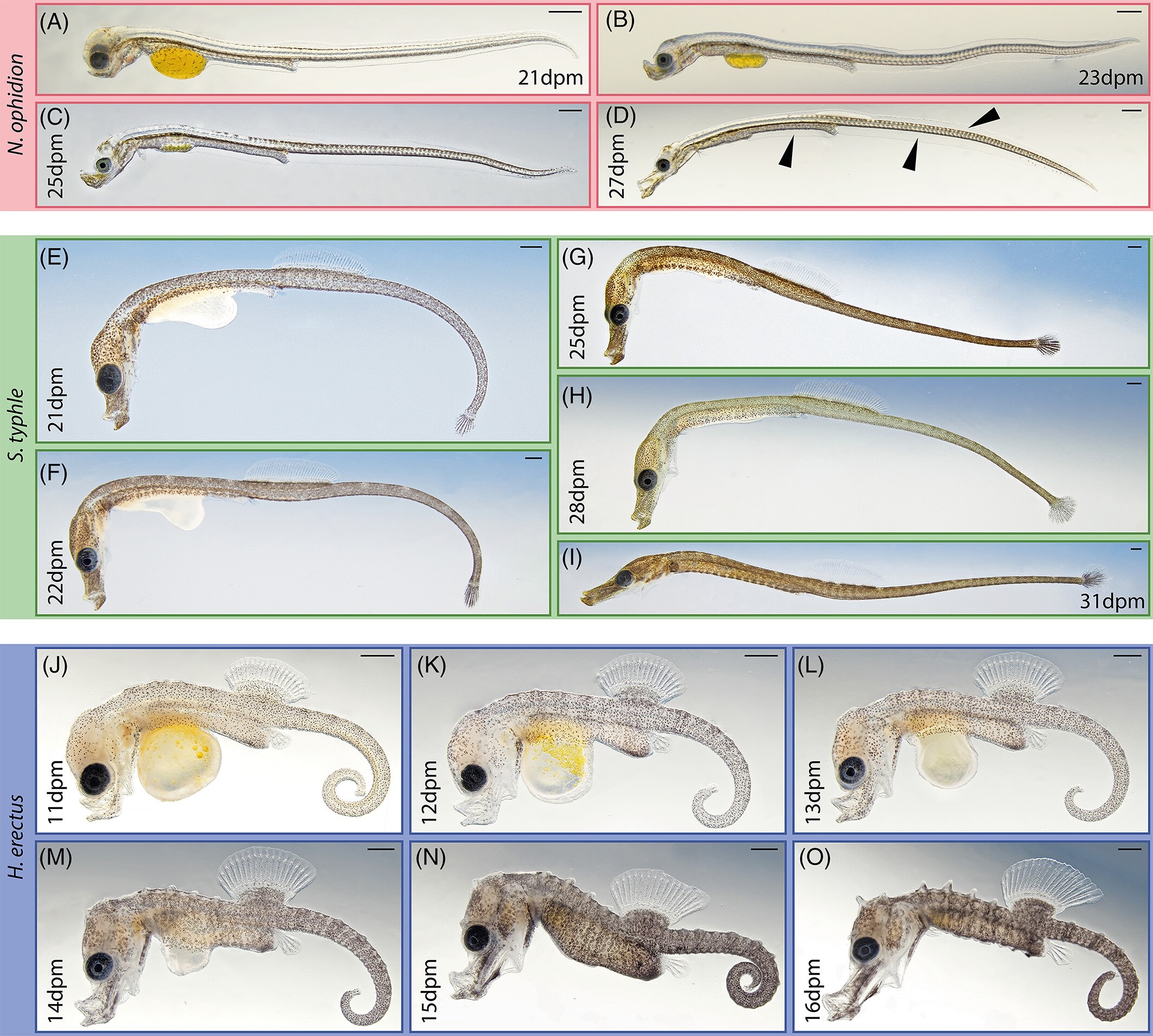
Organogenesis to release development in examined syngnathids. Nerophis ophidion (A-D), Syngnathus typhle (E-I), and Hippocampus erectus (J-O), respectively. The last prerelease period is characterized by snout elongation, continued pigmentation and the conclusion of allometric fin outgrowth. Arrowheads: black = fin fold. Scale is 500 μm; dpm = days post mating
And that’s where I see the problem with the paedomorphosis explanation. This is not simply a case of developmental arrest. There are clear differences in growth prior to the pharyngula stage, and the pharyngula stage is, at best, a point of divergence in development, and so much of what is happening at that point and thereafter is the appearance of evolutionary novelties. It’s not so much that the pattern stops, as that there are a whole host of additions to the organization of the syngnathid body plan in embryogenesis.
Also, data is always pretty.