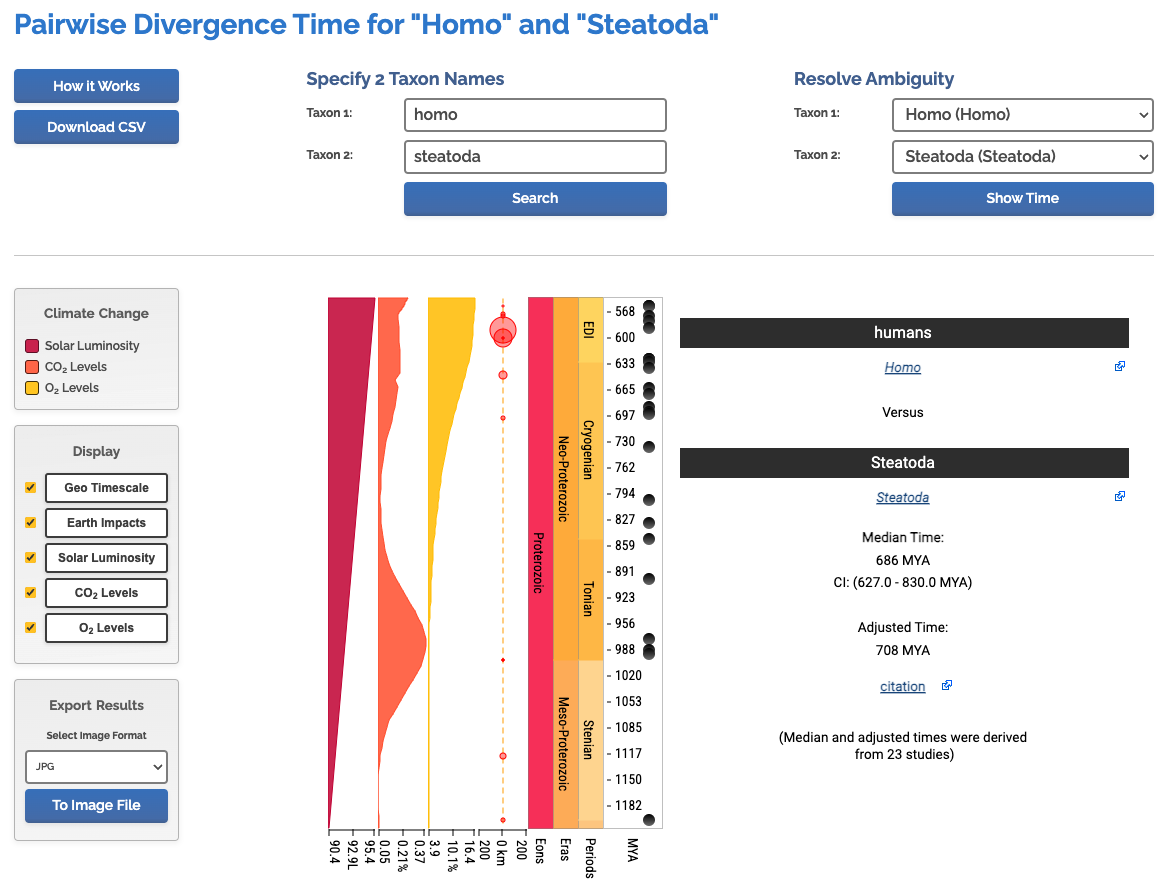
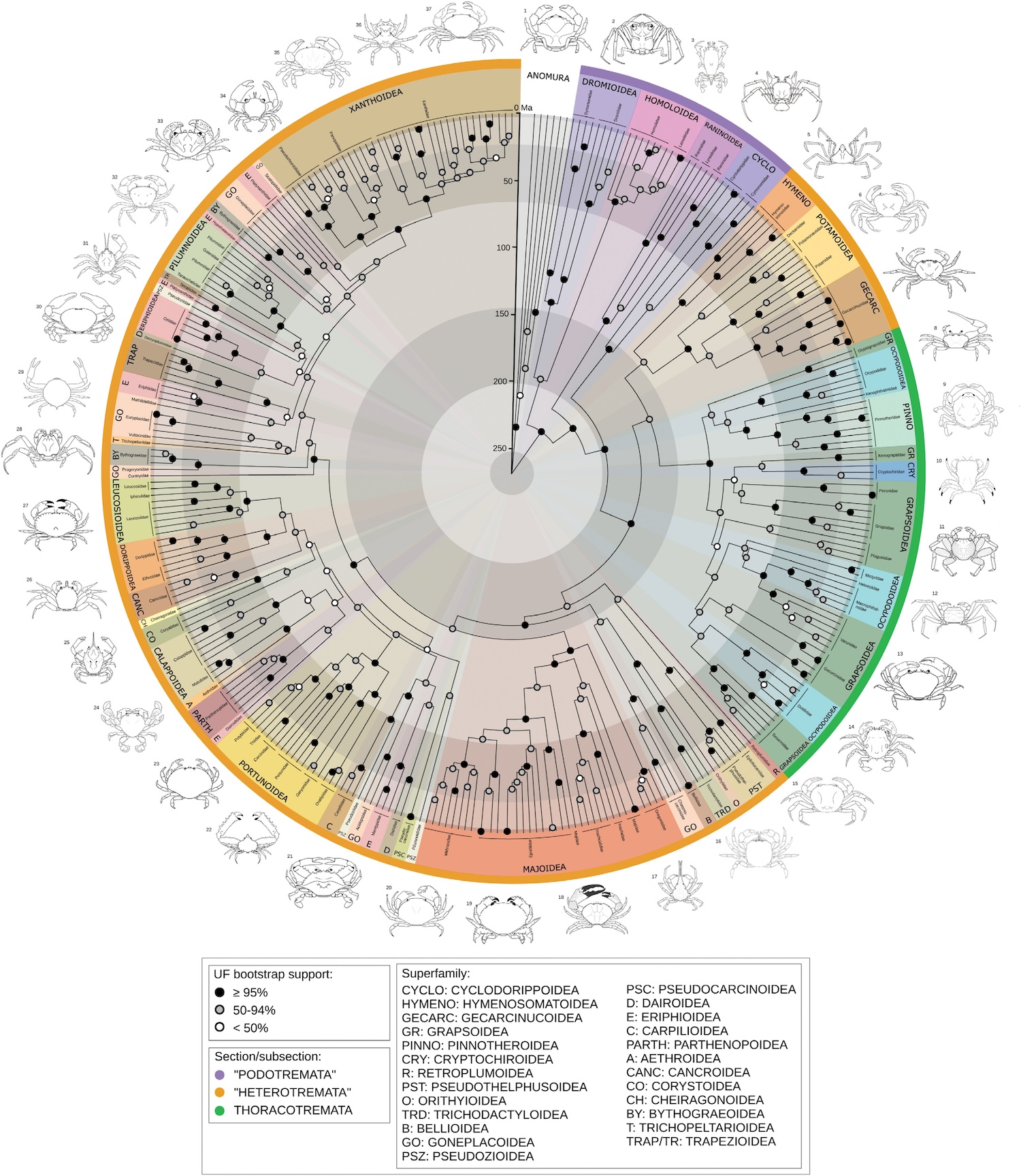
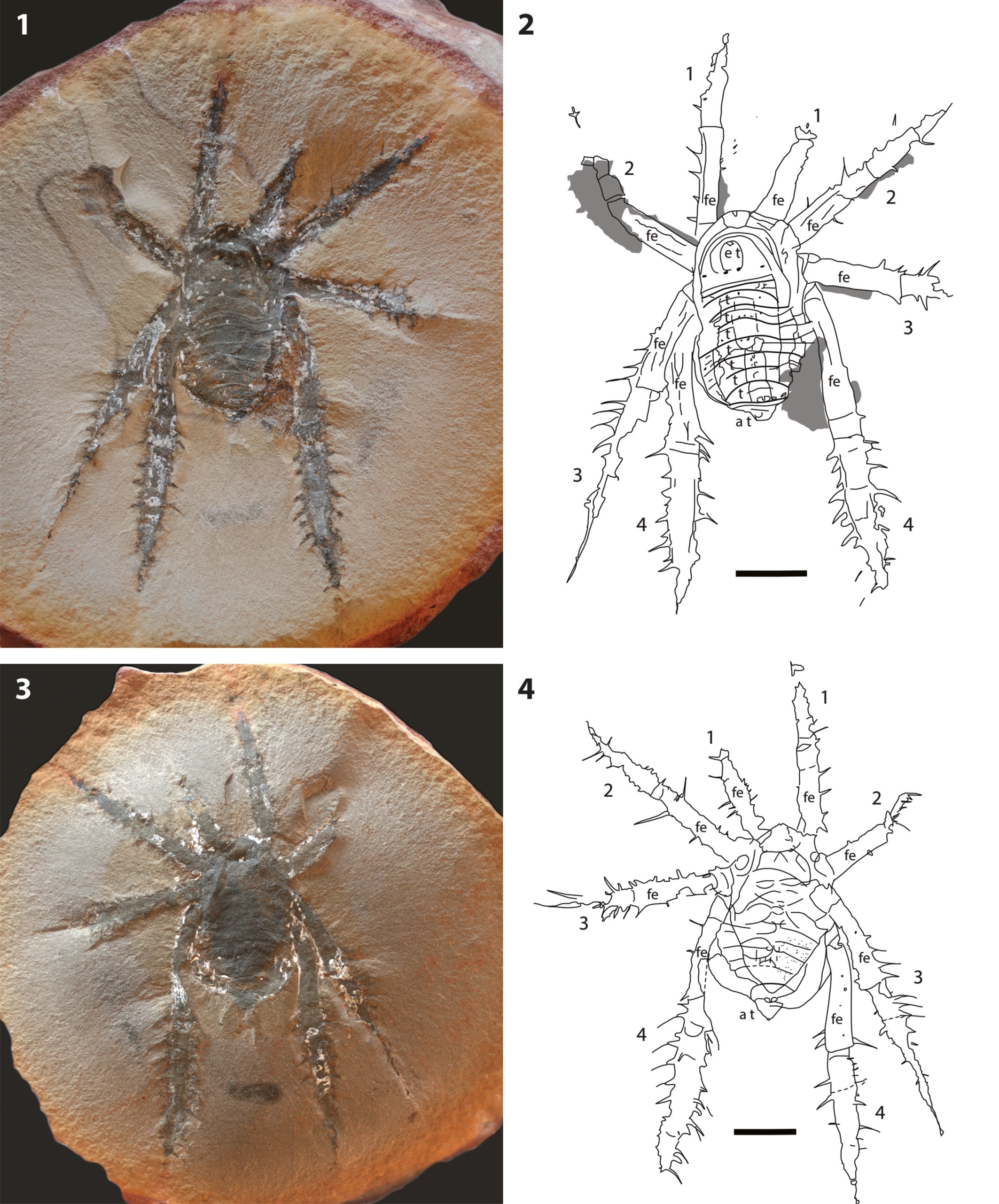
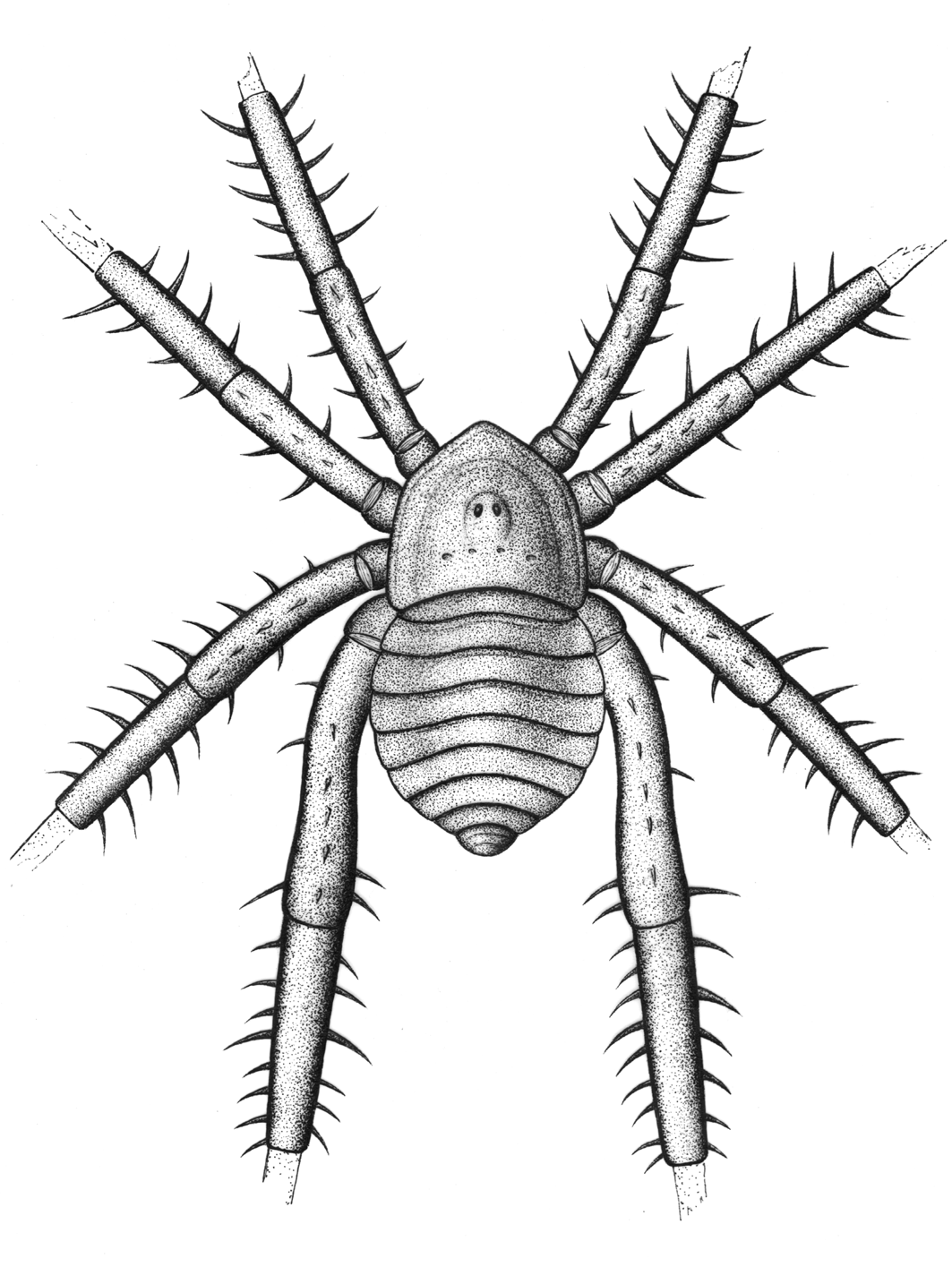
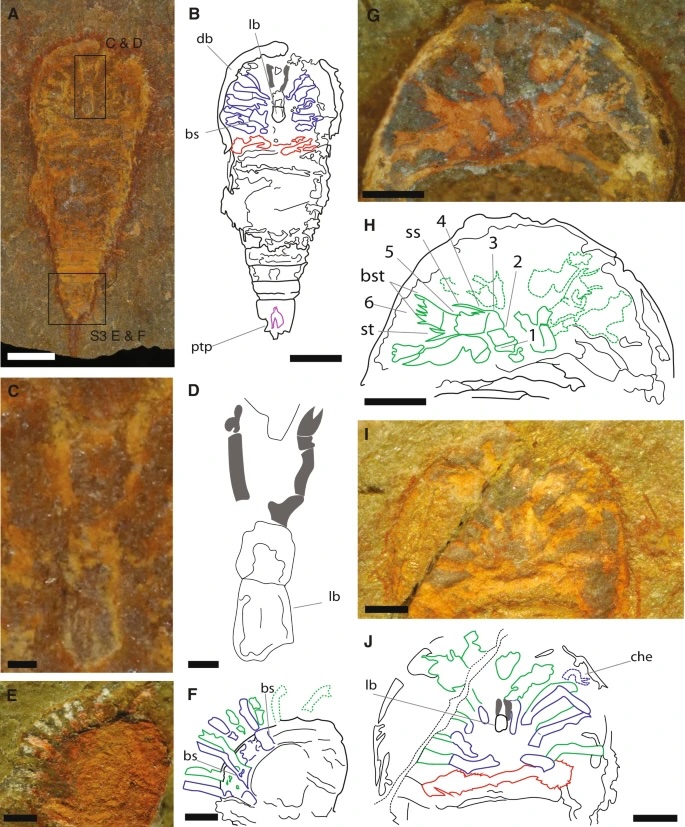
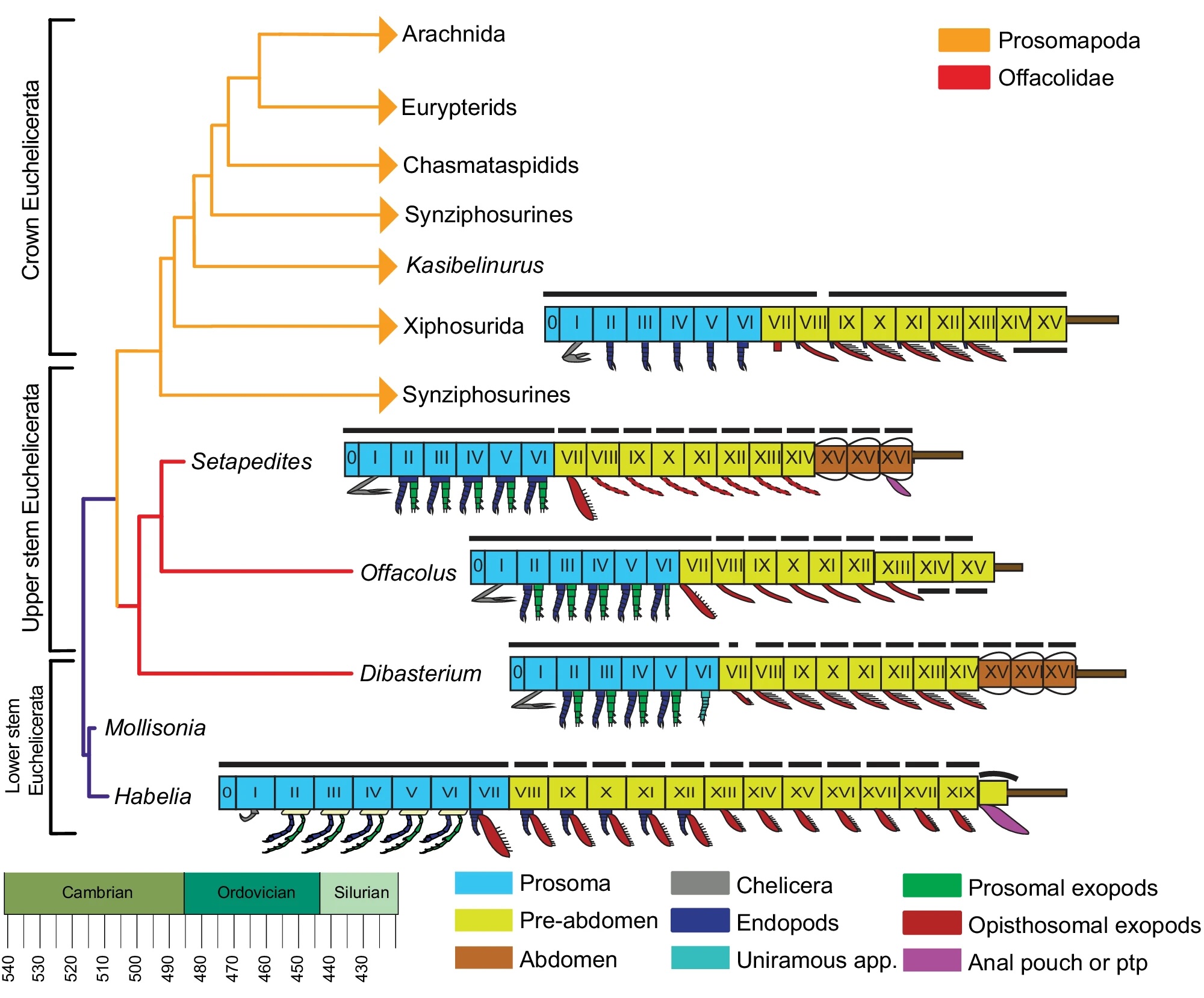
This is nice because it focuses mainly on deep evolutionary history, talking about how, contrary to common opinion, spiders are not particularly closely related to insects.
They’re all crunchies together, rather than squishies. Isn’t that enough to combine them in an evolutionary taxonomy?
(Nope.)