Dang. I’m not a botanist, and I’m honestly a bit weak on all that plant stuff, but I have to give some background on plant anatomy and photosynthesis to give some context for this cool story. Fortunately I was explaining all this to students in cell biology last week, so I can manage!
First, you all know that plants make sugar from carbon dioxide and sunlight. They use photosynthesis in a set of light reactions to produce energy (in the form of reducing compounds and ATP) that are passed on to a pathway called the Calvin cycle, which fixes CO2 into carbon compounds. It does that by adding the carbon in CO2 to a 5-carbon sugar called ribulose bisphosphate, producing a 6-carbon molecule that is immediately split into two 3-carbon molecules, called 3-phosphoglycerate or 3PG. The enzyme that carries out this reaction is called rubisco, and it’s not particularly efficient. In fact, it’s kind of terrible — it works poorly in a low CO2 environment (like modern Earth!), and plants can lose 25% of their energy to a reaction with O2, rather than CO2. The Calvin cycle is thoroughly intertwined with all kinds of reactions in plant biochemistry, though, so it’s pretty much indispensible. There isn’t an alternative, more efficient reaction that can substitute for it.
Evolution has been creating workarounds, though! Some plants have evolved a kind of supercharger for CO2 — they use an alternative enzyme, PEP carboxylase, to fix CO2, adding the carbon to a 3-carbon intermediate, phosphoenol pyruvate, to produce a 4-carbon molecule, oxaloacetate, which is then passed along to other cells where the carbon is cleaved off to form CO2 again, which sounds kind of pointless, I know…except that what it does is create a CO2-rich environment in the destination cells, so rubisco can run much more efficiently. See? A turbocharger for plant sugar synthesis.
These plants also have a specific anatomical organization, called the Kranz (German for wreath) pattern. There is an outer ring of mesophyll cells that specialize in fixing carbon with PEP carboxylase, and they transport the 4-carbon intermediate into an inner ring of cells, the bundle sheath cells, where rubisco re-fixes the CO2 into a 3-carbon intermediate.
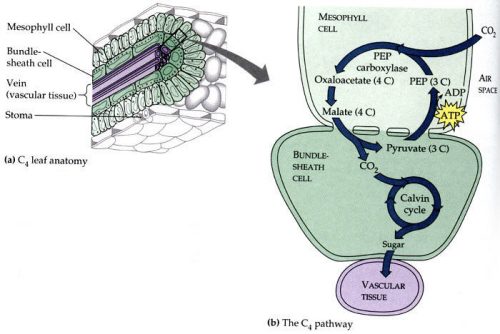
Not all plants have this ability. The plants that don’t, that rely entirely on just the bare bones Calvin cycle that produces a 3-carbon intermediate, are called C3 plants. Familiar C3 plants are wheat, rice, and barley. The plants that do have a supercharger and produce a 4-carbon intermediate are called C4 plants. Corn and sugar cane are well-known C4 crops. C4 is better at coping with environments poor in CO2, like everywhere. What if we could transplant that C4 metabolism in crop plants that lack it, like wheat and rice? We’d expect significant improvements in growth.
You might argue against that by noting that the Kranz anatomy is rather specific and detailed…but it turns out that Kranz anatomy is not essential for terrestrial C4 plant photosynthesis. Some plants have the C4 enzymes without the mesophyll/bundle sheath cell arrangement, and they benefit. It may also be feasible to engineer a proto-Kranz arrangement into C3 plants as a first step, and this is being done:
The C4 photosynthetic pathway accounts for ∼25% of primary productivity on the planet despite being used by only 3% of species. Because C4 plants are higher yielding than C3 plants, efforts are underway to introduce the C4 pathway into the C3 crop rice. This is an ambitious endeavor; however, the C4 pathway evolved from C3 on multiple independent occasions over the last 30 million years, and steps along the trajectory are evident in extant species. One approach toward engineering C4 rice is to recapitulate this trajectory, one of the first steps of which was a change in leaf anatomy. The transition from C3 to so-called “proto-Kranz” anatomy requires an increase in organelle volume in sheath cells surrounding leaf veins. Here we induced chloroplast and mitochondrial development in rice vascular sheath cells through constitutive expression of maize GOLDEN2-LIKE genes. Increased organelle volume was accompanied by the accumulation of photosynthetic enzymes and by increased intercellular connections. This suite of traits reflects that seen in “proto-Kranz” species, and, as such, a key step toward engineering C4 rice has been achieved.
Key things to note: they are recapitulating known evolutionary pathways to more rapidly ‘evolve’ a C3 plant to a C4 state. They’ve generated a line of rice with the first step in this pathway, the proto-Kranz condition. This does not, however, mean that they’ve produced a rice plant with higher yields — they have yet to introduce all the other steps in C4 metabolism. They do state that this transition, while not increasing efficiency yet, has also not reduced the yield of the rice plant, which suggests that the initial steps in the evolution of this pathway did not involve a cost to the plant, and also that the morphological changes, which I would have naively thought would be the biggest obstacle, may have been relatively trivial.
The complexity of the anatomical and biochemical changes needed for the C3-to-C4 transition appears seemingly incongruent with the multiple independent origins of the pathway. However, the results presented here suggest that one of the earliest steps in C4 evolution, the transition from C3 to protoKranz, could have resulted from modified activity of a single gene.
Now onward, to radically engineered biological organisms!
Voznesenskaya EV, Franceschi VR, Kiirats O, Freitag H, Edwards GE. (2001) Kranz anatomy is not essential for terrestrial C4 plant photosynthesis. Nature 414(6863):543-6.
Wang P, Khoshravesh R, Karki S, Tapia R, Balahadia CP, Bandyopadhyay A, Quick WP, Furbank R, Sage TL, Langdale JA (2017) Re-creation of a Key Step in the Evolutionary Switch from C3 to C4 Leaf Anatomy. Curr Biol. doi: 10.1016/j.cub.2017.09.040

That is seriously cool.
Comparing ancestral teosinte to modern maize, I sort of wonder if future GM rice will have grains the size of peas. Or would that not be the way C4 rice would work? Anyone who does know–please, in your explanation, remember that I am utterly utterly ignorant, so use small words.
Cuttlefish – It wouldn’t necessarily mean bigger grains, just because the plant is producing more energy doesn’t automatically mean it’s grains will be bigger; it could mean more grains per plant or the plant maturing more quickly.
As I am pretty sure I understood at least one word in three (thank heavens for articles and conjunctions) this looks rather impressive. Another green revolution?
This. Is. Rad.
Now watch Greenpeace and similar organisations misunderstand and oppose this.
Or we could just pump a lot more carbon dioxide into the atmosphere. I’m sure that wouldn’t have any unintended consequences.
Every now and then you read a science article that doesn’t make you feel like a colossal dumb shit. This gives me actual hope for the future as well.
Yep. Knew that.
Knew everything but the name “Calvin cycle”, though I’m sure I was taught that at some point. No knock on my teachers, but molecular biology was never my thing. I’m more likely to remember details of cladograms than names of molecular pathways.
Um, no. Did not know that. Probably never knew that.
I guess I’ll just take you word for it – you said these are the basics? Okay then. Guess I have 5000 pages of reading to do soon.
But at least I understand the significance of the conclusion:
So, if I understand correctly, there was concern that the increased organelle size and – I’m guessing here – the relationship of nutrient transfer mechanisms (like stomata) to the locations where nutrients are used (mesenchymal cells in c4 plants) would create inefficiencies that might only persist in a population given specific selection pressures (absence or presence of an environmental selection pressure that would otherwise make the trait highly disadvantageous).
Although with further refinement of nutrient transfer mechanisms the latter part might be overcome, the piece about organelle size (and something else I missed?) was expected to have non-neutral and negative effects unless and until the c4 process got going. This would not only make it difficult to explain how c4 managed to take hold in multiple different lineages, but also would foreshadow a significant hurdle in modifying c3 plants for advantages in human agriculture through a fully realized c4 photosynthetic system.
Instead, however, the researchers found that inducing the primitive structural state required to eventually realize c4 photosynthesis required less intervention than was (thought? feared?) and/or the primitive “proto-Kranz” morphologies induced far less negative efficiency than thought, with the researchers not able to actually detect negative efficiency at this point in their studies, so the effect is likely to be small if it is present at all.
The near-term practical importance of this would seem to be that it is easier to engineer/breed plants with the proto-Kranz condition for further work in inducing the rest of the c4 cycle and the longer-term practical importance is that plant varieties can be replaced in stages, with proto-Kranz varieties spread even before research on fully modifying a plant for c4 photosynthesis is completed. Because of this, it might (though we can’t yet know) be possible to spread the final modifications sooner and easier both because the widely-dispersed populations of proto-Kranz varieties might have an easier time taking up genes by reproducing with engineered c4 plants but also because the whole research process might be finished much earlier given that what was thought to have required multiple steps in fact requires fewer, perhaps only one.
Have I got this right?
C3 and C4 plants have different δ13C isotopic signatures. Both types prefer the lighter 12C, however, C4 plants are less picky. I wonder if this proto-Kranz anatomy would generate a measurable change in isotope affinity.
If I recall correctly, C4 photosynthesis also requires less *water* than C3. So the success of the C4-rice project might not only improve yields, but also convert rice from a water-intensive crop to a drought-tolerant one.
So awesome, in agribiotech starting on Monday, will so use this!
DonDueed: let’s run a double-blind randomized control trial to check. Oh wait.
this kind of bio-engineering sounds reasonable and may yield helpful results with fewer negative side effects as opposed to making crops immune to weed killers enabling the profligate use of same with predictable and unpredictable results.
uncle frogy
Since C4 plants have not completely replaced C3 plants, the latter must have some selective advantage(s). Is it known what this/these is/are? If the C4 pathway had only evolved once, it could be the case that the plants it evolved in had simply not had time to diversify into all plant niches. But since:
that seems doubtful.
If C4 plants come from C3 plants, why are there still C3 plants?
KG it may simly be that the C4 pathway didn’t give a sufficient selective advantage to out compete the existing C3s, just enough to carve a niche in already crowded ecosystems. However I seem to remember that C4 is better at taking up CO2 at higher temperatures, one of the downsides of climate change being that contrary to what one might expect a lot of crop yields will drop as temperature increases, so altering C3 pathways to C4 pathways could be extremely important in keeping people fed while as it gets hotter.
Last reply also relevant to rietplum.
@Jazzlet:
rietplum was engaging in snark in the longstanding Pharyngula tradition of, “If humans are descended from monkeys, why are there still monkeys?” which originated as a non-snarky question asked by creationists who think they’re being clever.
@ Crip Dyke etc
Uff, not enough coffee, obvious now you’ve pointed it out.
More Intelligent Design™, izzit? What a Rube Goldberg method of improving productivity! And then the Intelligent Designer gave his Chosen People™ one of the less productive crops as a staple. Why, one might almost think no one’s minding the store….
Huh, I was out for a while and suddenly my professional field is being discussed!
Water use and carbon fixation in plants are deeply intertwined, because plants mainly use water to replace transpiration from their leaves, which depends on stomatal opening. Plants can control water loss by closing their stomata (tiny air holes on leaves) but this cuts off mesophyll ventilation, resulting in CO2 shortage. This is the main reason why even mild drought cuts plant productivity, and why drought resistant crops can’t be highly productive in drought conditions although they can survive better than non-resistant crops.
C4 plants can manage with lower mesophyll CO2 levels so they need less stomatal opening/mesophyll ventilation, so they use less water per amount of carbon fixed. Obviously, higher water use efficiency is useful in nearly all growing conditions. Even a wetland crop like rice might benefit from this, if water transport from roots to leaves is a limiting factor on hot sunny days. More importantly, if I understand correctly, C4 improves the basic capacity of photosynthetic machinery, which allows a very high output if both water and light are abundantly available.
KG:
Good question. I don’t know but I might guess that the extra C4 cycle has a non-negligible energy cost.
jazzlet:
It is usually stated that C4 plants are most competitive in warm climates, and especially in sunny habitats. If I understand correctly, bright daylight can easily provide more energy than a C3 photosynthetic machinery can utilize, especially when CO2 intake is limited by water stress. The combination of bright light and limited water availability is typically associated with high temperatures, but I’m not sure if C4 success has anything to do with temperature as such.
It’s not that C4 is many times more productive than C3, but relatively few C4 species have a high presence in certain environments where C4 is competitive – and these are generally highly productive environments. Notably, these include most farming environments.
Rising atmospheric CO2 concentration may reduce the competitivity of C4 vs. C3 plants, but probably not dramatically, unless we really screw up the atmosphere.
Crip Dyke:
There must be some misunderstanding on either your part or mine. New modifications in cultivated plants are spread in farming environments by propagating new plant cultivars and planting them instead of older cultivars. I don’t think anyone wants to turn wild C3 plants into C4.
Now, it seems that engineering C4 cultivars for rice and other C3 crop plants is indeed very likely possible in near future.