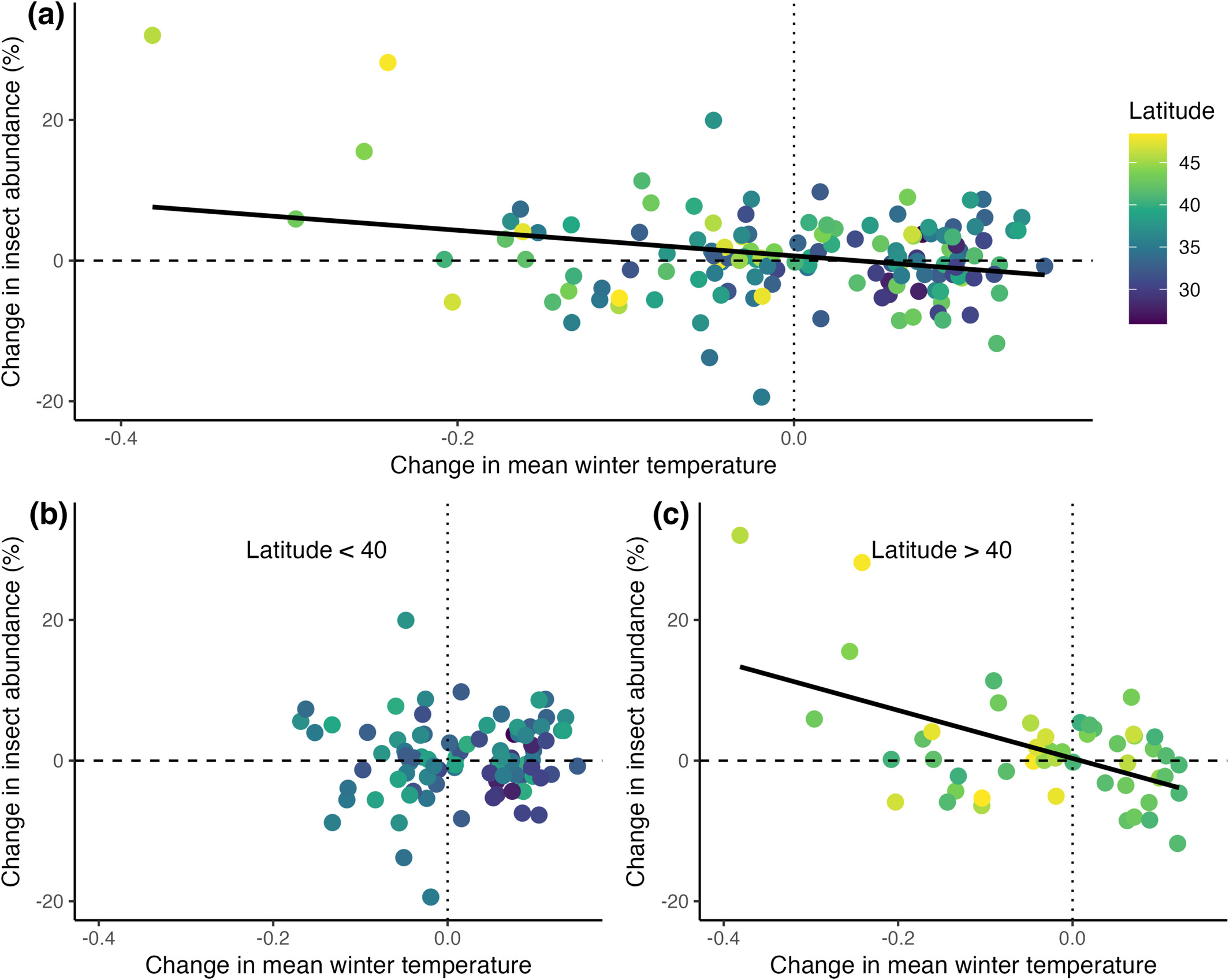
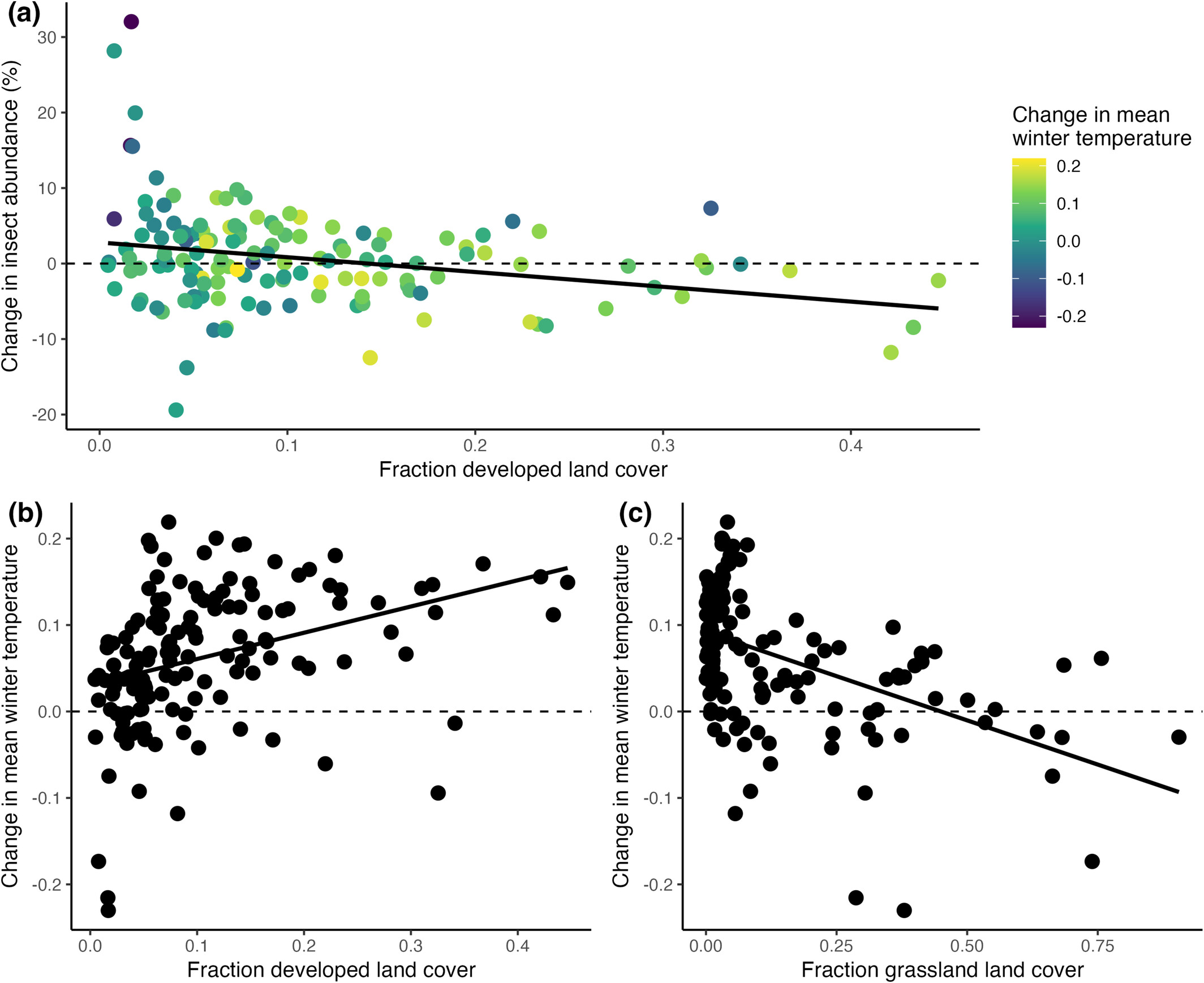
I think I’m trapped at home today — I tried walking to work, and didn’t get beyond my driveway, because we had a thaw and a refreeze and it’s slick as snot out there. Then we’re supposed to get more snow this morning, with temperatures plummeting down to -15°C with 50mph wind gusts, so I’m cowering at home today. The spiders will go hungry for a day (they are opportunistic feeders, they can handle it).
If you’re similarly stuck at home, here’s an hour long video that I thought was very good. It rips into a couple of self-styled “science” based influencers who are anything but.
The most shocking bit was seeing Joe Rogan getting furious at any push-back on his anti-vax views, and basically shutting down the conversation by claiming that the polio epidemic was co-incident with they years of heaviest DDT use. He also made the standard skeptical claim that vaccines were a late response to an already fading plague, which is sort of true. There are multiple approaches to a serious disease: behavioral shifts, like self-quarantine, and improved hygiene can reduce the incidence and severity of infections, but it takes efficacious medical responses to deliver the coup de grace. And Joe Rogan doesn’t understand science at all if he falls for the correlation equals causation canard. DDT does not cause polio.
The video also jumps on Bill Maher. He’s got this canned response to any claims, saying that we don’t know 100% of everything, more like 20% or 10%, so his weird fads might be true. It’s nonsense. Of course there is much left to learn, but we can say with 100% confidence that you shouldn’t eat cyanide, or that the earth is spherical, or that vaccines don’t cause autism, because smart, skeptical people have studied that stuff and have objective data to back up their arguments. We don’t even quantify knowledge as a percentage fraction of everything, so that’s a bogus metric anyway. I’m willing to go along with a claim that we only know 0.00001% of everything, but that the bits we know, we know pretty damn well, so please, Bill Maher, don’t jump off the roof of a New York skyscraper to test your ‘theory’ of gravity.
Another good topic was about what having a PhD means. It’s not a free pass to make everything you say valuable, important, and true. It just says you passed an apprenticeship. You presumably got some training in critical thinking which the Joe Rogans of the world lack, but you have to demonstrate your skills throughout your life. There are also some really bad theses out there — there is some pressure to get students out the door so you can get a new crop started, and some bad PIs who will let garbage pass as long as they get a publication out of it.
(By the way, I think my PhD thesis holds up. Not only did multiple researchers build on it afterwards, but it wasn’t even just mine — it was the product of a collaboration with several absolutely brilliant mentors and colleagues, which is how every thesis ought to be.)