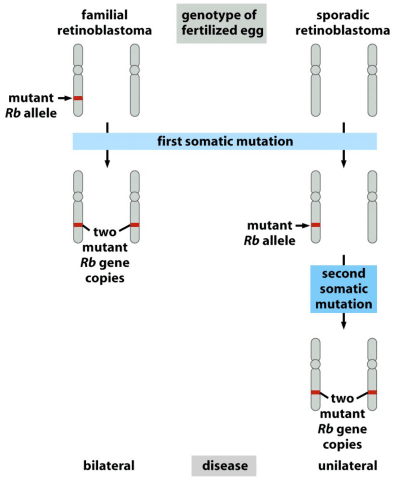
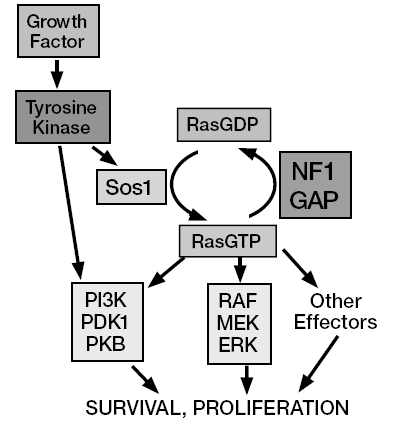
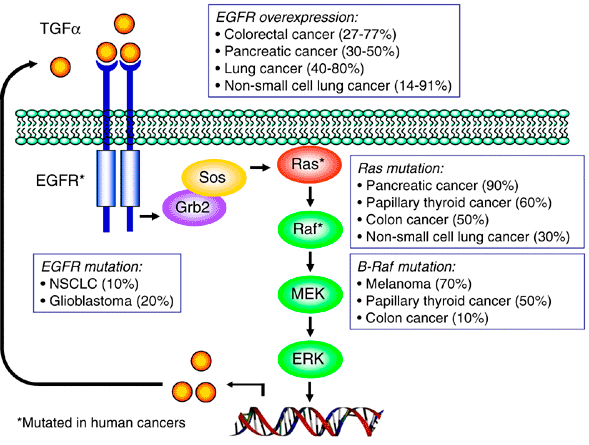
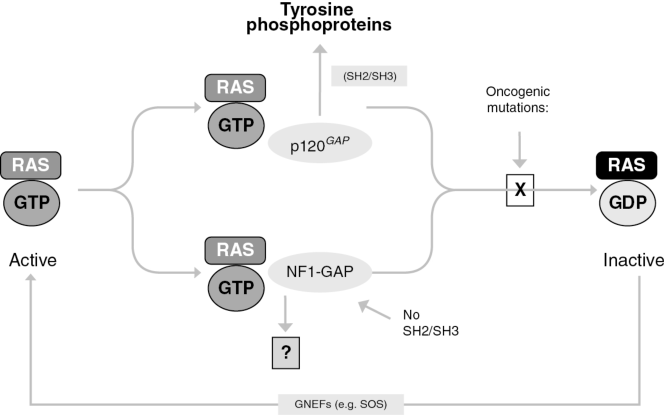
You go, guys, but…what can I do to support a charity fundraiser for men’s health issues when the gimmick is to grow a moustache? It’s not as if I can grow a second one. I guess all I can do is urge all of you poor barefaced men to join Movember, and let your face do its manly thing.
But then, there’s the dilemma — I have to tell you to not shave it off in December, so you won’t be able to do it again. What’s with all these guys with naked lips, anyway? Don’t you get cold? How do you filter plankton?