It’s been a while since I brought everyone up to date on the progress of my Ecological Development course, because I’ve been busy. So have the students. After our spring break I subjected them to the dreaded oral exam, which actually isn’t so bad. I tried to engage them less in an adversarial role and more as a quiet conversation between two people on science. Some students took to it easily — the more outgoing ones — others were noticeably nervous, which was OK, and I hope they learned that it isn’t that terrifying to have a discussion with a mentor.
Then the next few weeks were a mad whirl of horrible things done to babies: teratogens, endocrine disruptors, multi-generational epigenetic inheritance, all that fun stuff. We wrapped up with the depressing stuff for me, although the young’uns were more sanguine, I think. We talked about adult onset developmental diseases (it’s good to look at heart disease through the lens of developmental biology), aging, and cancer. Next week they get some time off, because I’m being drawn away to a conference on the east coast, but they’re supposed to spend it preparing for their final presentations, which will consume the last two weeks of class. And then we’re all done. School’s out for summer!
I’m going to say a bit about our last class discussion, because it got into some interesting territory and reflects the theme of the course well. We talked about that theory mentioned in the title of this article, and the origins of cancer, and to do that I have to give you all a little background.
Tissue Organization Field Theory (TOFT) is an alternative to what is sort of the dominant paradigm in cancer biology, the Somatic Mutation Theory (SMT). I have to say “sort of” because what I get from the literature is that SMT is more of a working assumption, and that cancer biologists are open to new ideas. The SMT is a useful molecular perspective on carcinogenesis. It postulates that cancer is a cellular disorder in which the genetic material has been perturbed to produce a lineage of cells with aberrant characteristics, that if we want to figure out what the primary cause of a cancer was, we can trace it back to a somatic mutation, or a change in a critical gene or, more likely, multiple genes, that lead to uncontrolled proliferation. So we pursue oncogenes, genes that have the potential to acquire mutations that trigger cell division or bypass control points, and tumor suppressor genes, protective genes that, when damaged, remove essential regulators of growth.
So, under the SMT, cancer is a disease caused by the progressive accumulation of mutations in cells of the body, as they divide. These mutations gradually strip away the normal restraints on cell division, and on immune system recognition, and on cell death activation, etc., etc., etc. until you have a rogue cell that can seed the growth of a massively disruptive tumor.
And it’s not wrong! Cancer biology has been immensely productive in identifying the enabling mutations, and even developing treatments that specifically target molecular agents of cancer. We know that somatic mutations are a routine part of the progression of cancer, and we also know that there heritable alleles that can affect the likelihood of the disease. The SMT is a tool to explain many of the phenomena of cancer, and it’s not going to just go away. It’s also a tool that is amenable to a reductionist approach to cancer biology, and is well-adapted to the utility of molecular biology.
Tissue Organization Field Theory is an alternative explanation for the origins of cancer.
TOFT argues that the focus of the SMT on single cell events is inappropriate and misses a whole range of effects at the level of tissue organization, effects which are more important in creating a pathological environment in which those mutations can accumulate. Further, it gets into field theory, which is important in developmental biology but isn’t exactly the subject of common conversation. Here’s one standard definition of a field: “a morphogenetic (or developmental) field is a region or a part of the embryo which responds as a coordinated unit to embryonic induction and results in complex or multiple anatomic structures.” If that’s not helpful — and it probably isn’t, we’d have to go over a textbook if we wanted to explain developmental field theory — here’s a diagramatic metaphor. Do you see the field in this picture?

There’s something special about part of that image, but it’s not that the individual subunits are intrinsically different — it’s tied up in the relationships between the central set of blocks and the blocks outside of it. There’s something different going on with a subset of the blocks, but it’s not necessarily best described by explaining the details of single blocks, but is more easily explained at a higher level, as properties of a tissue within a tissue. Of course, what will eventually happen in a developing organism is that those central blocks will express a unique pattern of genes, so eventually it’s identifiable by molecular markers, but the field first arises in a sea of genetically and epigenetically uniform cells.

Another important property of a field is that it is not itself uniform. It’s going to acquire complex spatial properties over time. Insect limbs, for instance, arise from a disc-shaped field with extensive patterning information within them, so the central region will become the distal tip of the limb, and there is information that is interpreted as polar coordinates that specifies what portion of the limb is anterior, posterior, medial, and lateral (the limb is not a uniform cylinder). Similarly, vertebrates have a limb field represented in the limb bud, with gradients of morphogens specifying the orientation of the limb, and with re-expression of Hox genes used to specify longitudinal positions. Hox genes in a limb field are interpreted in different ways than Hox genes along the body axis, obviously.
The key factor here is that in field theory cells are not simply independent units — they are part of a larger assemblage, a tissue, that has complex fates that are not easily summarized by individual gene expression. They have to be understood as a network.
That’s the first thing to remember: TOFT is treating a cancer as a field, with field properties, which are not adequately described if you only look at cancer as a collection of autonomous cells all doing their own thing at the command of their broken genes. Aberrant disruption of the field can produce aberrant structures without requiring any genetic changes.
This is the difference between a mutagen and a teratogen. The effects of a mutagen are caused directly by damage to the structure or sequence of DNA; they produce heritable changes to the cells of an organism. Teratogens, on the other hand, are not necessarily mutagenic at all — they disrupt the normal pattern of development without changing genes at all. Thalidomide babies, for example, had some extreme morphological changes, like phocomelia or truncated limb development, but those are not heritable, and the people affected by thalidomide can grow up to have normal, healthy children.
TOFT argues something similar, that there is a disruption of a tissue that initializes aberrant growth, that may then be an enabling precondition for the accumulation of mutations. One piece of evidence for this is a set of experiments on tissues, illustrated below.
Most cancers arise in epithelial tissues, like the sheets of cells that line glands or your organs, in large part because those are the cells that divide most frequently. These epithelial cells, also called parenchyma, do not typically grow in isolation, but on a substrated of connective tissue, extracellular matrix, and other cell types, called stroma. The stroma supports and signals the overlying epithelium, and vice versa, and together they make a coherent functional tissue.
The theory suggests that cancers can arise in epithelia by way of disruptions in signaling in the stroma. A carcinogen could distort the interactions between stroma and epithelium at the level of the stroma, and the epithelium then goes nuts and proliferates to produce a pre-cancerous mass.
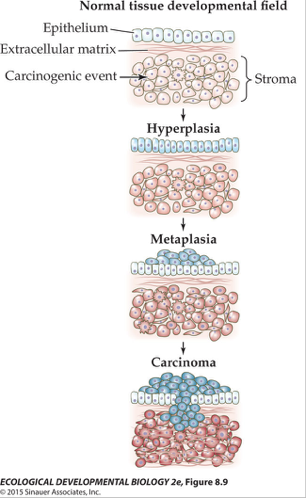
One test of the theory would be to separate stroma and epithelium, expose the stroma to a short-lived teratogen, and then after the teratogen was washed out, re-associate the two and determine whether there was an increase in the frequency of cancers in the epithelium, which has not been exposed to teratogens.
The experiment has been done. Here are the results for rat mammary gland tissue in which the epithelium was exposed to the solvent vehicle but no N-methyl nitorsourea (a potent mutagen), while the stroma was soaked in NMu, labeled VEH/NMu. The numerator describes the epithelial condition, and the denominator is the stroma condition, so NMu/NMu means both were hit with the mutagen, VEH/VEH means both were exposed only to the vehicle, and NMu/VEH means the epithelium was poisoned with NMu, while the stroma was not.

There’s an awfully strong positive correlation between exposing the stroma to mutagens and getting tumors, and a negative correlation with exposing epithelia to mutagens and tumors.
You want more evidence? Here’s a very interesting experiment. Start with aggressively metastatic melanoma cells from a human patient (labeled in green, below). Inject them into a completely different environment, the neural crest pathway in a developing chick embryo. Surprisingly, if you accept the SMT, the cancer cells calm right down and are conditioned by their environment to participate in normal development in the chick and get incorporated into the facial cartilages and sympathetic ganglia.

I suspect those melanoma cells do carry somatic mutations, and are not actually “cured” of a predisposition to cancer. What the experiment says, though, is that environmental influences are extremely important in regulating the behavior of these cells, and that modifying the cells communicating with the cancerous cells can have a profound effect on how they act.
Note that this is not a pathway to a cure. It’s all well and good to say that if we could break up a tumor, separate the individual cells and put them in a more nurturing, embryo-like environment, they’ll stop acting up and resume normal, regulated growth, but if we could do that, slicing out the tumor and tossing it in an incinerator would also be effective. The problem is that in a human patient we do not and cannot have such precise control of the micro-environment of the cancer, and in fact, the tumor itself is a kind of bubble of micro-environment that actively reinforces cancer growth.
My students and I read a paper from Carlos Sonnenschein, who is a major proponent of TOFT, as well as our textbook summary. The paper was titled The tissue organization field theory of cancer: a testable replacement for the somatic mutation theory. They’re a smart bunch, and they see the promise of the idea, in part because this whole course is about thinking a level above reductionist cell biology, but they also found the word “replacement” off-putting. It doesn’t invalidate everything about the SMT, but it does support an important alternative route for carcinogenesis. They also weren’t impressed by the rather aggressive insistence by some TOFT proponents that they have the One True Explanation, and that their observations are sufficent to explain cancer — we came up with a few alternative interpretations of their own favorite experiments that they haven’t nailed down completely just yet.
One thing that amused me is that the class consensus actually converged on the views of another paper by Bedessem and Ruphy, which I did not assign them to read, largely because of its more philosophical argument (I’ve focused on empirical/experimental papers in the class). This is how I feel about it, too.
The building of a global model of carcinogenesis is one of modern biology’s greatest challenges. The traditional somatic mutation theory (SMT) is now supplemented by a new approach, called the Tissue Organization Field Theory (TOFT). According to TOFT, the original source of cancer is loss of tissue organization rather than genetic mutations. In this paper, we study the argumentative strategy used by the advocates of TOFT to impose their view. In particular, we criticize their claim of incompatibility used to justify the necessity to definitively reject SMT. First, we note that since it is difficult to build a non-ambiguous experimental demonstration of the superiority of TOFT, its partisans add epistemological and metaphysical arguments to the debate. This argumentative strategy allows them to defend the necessity of a paradigm shift, with TOFT superseding SMT. To do so, they introduce a notion of incompatibility, which they actually use as the Kuhnian notion of incommensurability. To justify this so-called incompatibility between the two theories of cancer, they move the debate to a metaphysical ground by assimilating the controversy to a fundamental opposition between reductionism and organicism. We show here that this argumentative strategy is specious, because it does not demonstrate clearly that TOFT is an organicist theory. Since it shares with SMT its vocabulary, its ontology and its methodology, it appears that a claim of incompatibility based on this metaphysical plan is not fully justified in the present state of the debate. We conclude that it is more cogent to argue that the two theories are compatible, both biologically and metaphysically. We propose to consider that TOFT and SMT describe two distinct and compatible causal pathways to carcinogenesis. This view is coherent with the existence of integrative approaches, and suggests that they have a higher epistemic value than the two theories taken separately.
Anyway, keep an eye open for more on the tissue organization field theory — there seems to be a fair bit of ongoing debate in the scientific literature about it. I’ll keep telling everyone cancer is a developmental disease, so you need more developmental biologists to study it. Or, alternatively, every cancer biologist is already a developmental biologist.
Bedessem B, Ruphy S. (2015) SMT or TOFT? How the two main theories of carcinogenesis are made (artificially) incompatible. Acta Biotheor. 63(3):257-67.
Soto AM, Sonnenschein C (2011) The tissue organization field theory of cancer: a testable replacement for the somatic mutation theory. Bioessays. 33(5):332-40.

Does this mean that TOFT could be an explanation for why tumor promoters can cause tumor growth even though many are not mutagenic?
Yes. There’s a curious disconnect between the words “mutagenic” and “carcinogenic”. Many people assume they’re virtually synonymous, but they aren’t — there’s overlap, but there are carcinogens that are not mutagenic.
Growth factors would be a good example.
Fascinating. I really liked the quote from Bedessem and Ruphy. It makes perfect sense to me that different cancers would be more one type than the other.
I really wonder, though, what kind of applications continued research along these lines might lead to. Might we learn about new ways to determine whether a cancer is likely to become dangerous, especially while it is still very small? Could we learn about new ways to prevent some cancers? Any practical applications are doubtless decades away, but hopefully the better understanding of cancers will help somewhere.
I suspect that, should any one full theory does come to fruition, it’ll still be a combination of the two competing theories.
Where a teratogens cause a change in the field, however pre-cancerous and cancerous tumors can also cause alterations in their field, such as via increasing angiogenesis, immune masking, suppression of inflammation, etc.
I suspect that we’ll not find quite as many causes of cancers as there are cancers, but a gradient of causes to type, possibly, even by tissue type.
For, it’d be extraordinarily difficult to explain melanomas and the link between UV exposure and melanoma via TOFT, but easily one can explain that cancer via SMT.
Any chance this theory will be unified with gravity? Any predictions about dark matter this can make?
Oh wait, wrong web site…
Anyway, this was a pretty amazing post. I knew up until now that cells and their incredibly complicated interactions result in some incredibly complex things that can’t be reduced down to DNA, but up until now I had thought that the whole of such interdependencies could never be figured out and how the body ultimately works is hopelessly beyond our ability to fully understand. Treating it as a field seems like a very physicist thing to do, but it also gives me hope that it CAN be abstracted into some useful understanding some day.
@Dark Jaguar #5, the field in this case is biochemical intracellular signalling.
https://en.wikipedia.org/wiki/Cell_signaling
Certain cells would behave in specific ways at certain chemical gradients of signals from their neighboring, related (and sometimes, unrelated) tissues.
For a practical example, which has real world applications, interest and still ongoing research, see this.
https://en.wikipedia.org/wiki/Angiogenesis#Tumor_angiogenesis
PZ, do you know which paper the diagrams you used were derived from? It looks like it comes from Maffini 2004 (http://jcs.biologists.org/content/117/8/1495.long)? If so, then the diagram is hugely misleading as the right two groups (the control groups) had no surgery and therefore had stroma, epithelium, and fat cells exposed, and not epithelium only as the graph implies.
The graph also drops out both the numbers (which were small — that positive control rate of 100% was on a sample size of only 6), and the error bars (which were not that impressive). And hides the fact that there was a huge bias in animal survival rates (they only measured the rate in this rats who survived surgery, naturally, but that makes me wonder why there were 13 survivors in group 1 but only 8 survivors in group 2, and only 6 survivors in the control group that didn’t even have surgery!) I would also like to know why so many of their rats died under simple surgery (removing the mammary fat pads should not give ~40% mortality rate!).
Jeez, when you said some of the TOFT proponents go a bit overboard, you weren’t joking. I think this excerpt will make your head explode:
From https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4643632/
(Not that this invalidates TOFT, but by gum it raises the hackles.)
Why does the phenomena of rogue waves come to mind? A simple wave formula should explain, but reality is much more complicated.
http://oceanservice.noaa.gov/facts/roguewaves.html