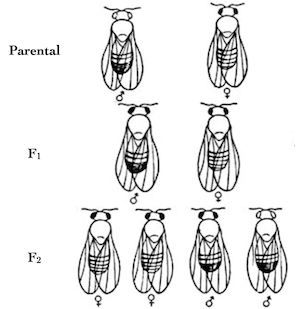
It’s the perfect storm, and I have myself to blame. I gave the fourth exam of the semester yesterday, and many of the students bombed. This happens every year, and it’s because the fourth exam is the hardest of them all: everything accumulates in this course, and it’s the combination of lots of math and understanding the topology of weird chromosomes entangled and swapping bits around that breaks their brains. Couple the fact that these are generally smart, high-achieving students with the shock of getting a 40% on an exam, and it’s going to get ugly. I’m going over the answers with them this afternoon.
Also, we have a major lab report due next week, and an in-class presentation. It is a high stress situation.
Students are already emailing me with requests for a point here and there on back assignments and exams. I’m going to spend my weekend with spreadsheets recalculating all their old assignments and putting together a final assessment, which will inspire a further frenzy of grade-grubbing.
If I drank anymore, this would be the weekend I’d finish off a bottle of good whisky, but I don’t drink at all anymore. I’d rather talk about genetics than grades, but the system will not allow that.