
Botanical Wednesday: I gotta be meeeeee!


I’ve been giving talks at scientific meetings on educational outreach — I’ve been telling the attendees that they ought to start blogs or in other ways make more of an effort to educate the public. I mentioned one successful result the other day, but we need more.
I give multiple reasons for scientists to do this. One is just general goodness: we need to educate a scientifically illiterate public. Of course, like all altruism, this isn’t really recommended out of simple kindness, but because the public ultimately holds the pursestrings, and science needs their understanding and support. Another reason, though, is personal. Scientific results get mangled in press releases and news accounts, so having the ability to directly correct misconceptions about your work ought to be powerfully attractive. Even worse, though, I tell them that creationists are actively distorting their work. This goes beyond simple ignorance and incomprehension into the malign world of actively lying about the science, and it happens more often than most people realize.
I have another painful example of deviousness of creationists. There’s a paper I’ve been meaning to write up for a little while, a Nature paper by David and Alm that reveals an ancient period of rapid gene expansion in the Archaean, approximately 3 billion years ago. Last night I thought I’d just take a quick look to see if anybody had already written it up, so I googled “Archaean genetic expansion,” and there it was: a couple of references to the paper itself, a news summary, one nice science summary, and…two creationist distortions of the paper, right there on the first page of google results. I told you! This happens all the time: if there’s a paper in one of the big journals that discusses more evidence for evolution, there is a creationist hack somewhere who’ll quickly write it up and lie about it. It’s a heck of a lot easier to summarize a paper if you don’t understand it, you see, so they’ve got an edge on us.
One of the creationist summaries is by an intelligent design creationist. He looks at the paper and claims it supports this silly idea called front-loading: the Designer seeded the Earth with creatures that carried a teleological evolutionary program, loading them up with genes at the beginning that would only find utility later. The unsurprising fact that many gene families are of ancient origin seems to him to confirm his weird idea of a designed source, when of course it does nothing of the kind, and fits quite well in an evolutionary history with no supernatural interventions at all.
The other creationist summary is from an old earth biblical creationist who tries to claim that “explosive increase in biochemical capabilities happened in anticipation of changes that were to take place in the environment”, a conclusion completely unsupportable from the paper, and also tries to telescope a long series of changes documented in the data into a single ancient event so that they can claim that the rate of innovation was so rapid that it contradicts the “evolutionary paradigm”.
So lets take a look at the actual paper. Does it defy evolutionary theory in any way? Does it actually make predictions that fit creationist models? The answer to both is a loud “NO”: it is a paper using methods of genomic analysis that produce evolutionary histories, it describes long periods of gradual modification of genomes, and it correlates genomic innovations with changes in the ancient environment. It is freakin’ bizarre that anyone can look at this work and think it supports creationism, but there you are, standard operating procedure in the fantasy world of the creationist mind.
Here’s the abstract, so you can get an idea of the conclusions the authors draw from the work.
The natural history of Precambrian life is still unknown because of the rarity of microbial fossils and biomarkers. However, the composition of modern-day genomes may bear imprints of ancient biogeochemical events. Here we use an explicit model of macro- evolution including gene birth, transfer, duplication and loss events to map the evolutionary history of 3,983 gene families across the three domains of life onto a geological timeline. Surprisingly, we find that a brief period of genetic innovation during the Archaean eon, which coincides with a rapid diversification of bacterial lineages, gave rise to 27% of major modern gene families. A functional analysis of genes born during this Archaean expan- sion reveals that they are likely to be involved in electron-transport and respiratory pathways. Genes arising after this expansion show increasing use of molecular oxygen (P=3.4 x 10-8) and redox- sensitive transition metals and compounds, which is consistent with an increasingly oxygenating biosphere.
This work is an analysis of the distribution of gene families in modern species. Gene families, if you’re unfamiliar with the term, are collections of genes that have similar sequences and usually similar functions that clearly arose by gene duplications. A classic example of a gene family are the globin genes, an array of very similar genes that produce proteins that are all involved in the transport of oxygen; they vary by, for instance, their affinity for oxygen, so there is a fetal hemoglobin which binds oxygen more avidly than adult hemoglobin, necessary so the fetus can extract oxygen from the mother’s circulatory system.
So, in this paper, David and Alm are just looking at genes that have multiple members that arose by gene duplication and divergence. They explicitly state that they excluded singleton genes, things called ORFans, which are unique genes within a lineage. That does mean that their results underestimate the production of novel genes in history, but it’s a small loss and one the authors are aware of.
If we were looking for evidence for evolution, we might as well stop here. The existence of gene families, for cryin’ out loud, is evidence for evolution. This paper is far beyond arguing about the truth of evolution — that’s taken for granted as the simple life’s breath of biology — but instead asks a more specific question: when did all of these genes arise? And they have a general method for estimating that.
Here’s how it works. If, for example, we have a gene family that is only found in animals, but not in fungi or plants or protists or bacteria, we can estimate the date of its appearance to a time shortly after the divergence of the animal clade from all those groups. If a gene family is found in plants and fungi and animals, but not in bacteria, we know it arose farther back in the past than the animal-only gene families, but not so far back as a time significantly predating the evolution of multicellularity.
Similarly, we can also look at gene losses. If a gene family or member of a gene family is present in the bacteria, and also found in animals, we can assume it is ancient in origin and common; but if that same family is missing in plants, we can detect a gene loss. Also, if the size of the gene family changes in different lineages, we can estimate rates of gene loss and gene duplication events.
I’ve given greatly simplified examples, but really, this is a non-trivial exercise, requiring comparisons of large quantities of data and also analysis from the perspective of the topologies of trees derived from that data. The end result is that each gene family can be assigned an estimated date of origin, and that further, we can estimate how rapidly new genes were evolving over time, and put it into a rather spectacular graph.
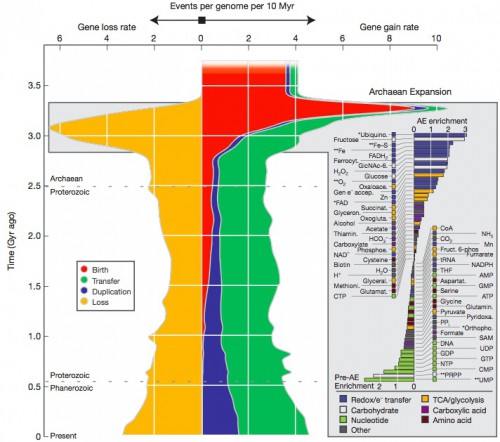
Look first at just the red areas. That’s a measure of the rate of novel gene formation, and it shows a distinct peak early in the history of life, around 3 billion years ago. 27% of our genes are very, very old, arising in this first early flowering. Similarly, there’s a slightly later peak of gene loss, the orange area. This represents a period of early exploration and experimentation, when the first crude versions of the genes we use now were formed, tested, discarded if inefficient, and honed if advantageous.
But then the generation of completely novel genes drops off to a low to nonexistent rate (but remember, this is an underestimate because ORFans aren’t counted). If you draw any conclusions from the graph, it’s that life on earth was essentially done generating new genes about one billion years ago…but we know that all the multicellular diversity visible to our eyes arose after that period. What gives?
That’s what the blue and green areas tell us. We live in a world now rich in genetic diversity, most of it in the bacterial genomes, and our morphological diversity isn’t a product so much of creating completely new genes, but of taking existing, well-tested and functional genes and duplicating them (blue) or shuffling them around to new lineages via horizontal gene transfer (green). This makes evolutionary sense. What will produce a quicker response to changing conditions, taking an existing circuit module off the shelf and repurposing it, or shaping a whole new module from scratch through random change and selection?
This diagram gives no comfort to creationists. Look at the scale; each of the squares in the chart represents a half billion years of time. The period of rapid bacterial cladogenesis that produced the early spike is between 3.3 and 2.9 billion years ago — this isn’t some brief, abrupt creation event, but a period of genetic tinkering sprawling over a period of time nearly equal to the entirety of the vertebrate fossil record of which we are so proud. And it’s ongoing! The big red spike only shows the initial period of recruitment of certain genetic sequences to fill specific biochemical roles — everything that follows testifies to 3 billion years of refinement and variation.
The paper takes another step. Which genes are most ancient, which are most recent? Can we correlate the appearance of genetic functions to known changes in the ancient environment?
the metabolites specific to the Archaean Expansion (positive bars in Fig. 2 inset) include most of the compounds annotated as redox/e– transfer (blue bars), with Fe-S-binding, Fe-binding and O
2-binding gene families showing the most significant enrichment (false discovery rate<5%, Fisher’s exact test). Gene families that use ubiquinone and FAD (key metabolites in respiration pathways) are also enriched, albeit at slightly lower significance levels (false discovery rate<10%). The ubiquitous NADH and NADPH are a notable exception to this trend and seem to have had a function early in life history. By contrast, enzymes linked to nucleotides (green bars) showed strong enrichment in genes of more ancient origin than the expansion. The observed bias in metabolite use suggests that the Archaean Expansion was associated with an expansion in microbial respiratory and electron transport capabilities.
So there is a coherent pattern: genes involved in DNA/RNA are even older than the spike (vestiges of the RNA world, perhaps?), and most of the genes associated with the Archaean expansion are associated with cellular metabolism, that core of essential functions all extant living creatures share.
Were we done then, as the creationists would like to imply? No. The next major event in the planet’s history is called the Great Oxygenation Event, in which the fluorishing bacterial populations gradually changed the atmosphere, excreting more and more of that toxic gas, oxygen.
What happened next was a shift in the kinds of novel genes that appeared: these newer genes were involved in oxygen metabolism and taking advantage of the changing chemical constituents of the ocean.
Our metabolic analysis supports an increasingly oxygenated biosphere after the Archaean Expansion, because the fraction of proteins using oxygen gradually increased from the expansion to the present day. Further indirect evidence of increasing oxygen levels comes from compounds whose availability is sensitive to global redox potential. We observe significant increases over time in the use of the transition metals copper and molybdenum, which is in agreement with geochemical models of these metals’ solubility in increasingly oxidizing oceans and with molybdenum enrichments from black shales suggesting that molybdenum began accumulating in the oceans only after the Archaean eon16. Our prediction of a significant increase in nickel utilization accords with geochemical models that predict a tenfold increase in the concentration of dissolved nickel between the Proterozoic eon and the present day but conflicts with a recent analysis of banded iron formations that inferred monotonically decreasing maximum concentrations of dissolved nickel from the Archaean onwards. The abundance of enzymes using oxidized forms of nitrogen (N2O and NO3) also grows significantly over time, with one-third of nitrate-binding gene families appearing at the beginning of the expansion and three-quarters of nitrous-oxide-binding gene families appearing by the end of the expansion. The timing of these gene-family births provides phylogenomic evidence for an aerobic nitrogen cycle by the Late Archaean.
So I don’t get it. I don’t see how anyone can look at that diagram, with its record of truly ancient genomic changes and its evidence of the steady acquisition of new abilities correlated with changes in the environment of the planet, and declare that it supports a creation event or front-loading of biological potential in ancestral populations. That makes no sense. This is work that shouts “evolution” at every instant, yet some people want to pretend it’s an endorsement of theological hocus-pocus? Madness.
Scientists, you need to be aware of this. The David and Alm paper is an unambiguously evolutionary paper, using genomic data to describe evolutionary events via evolutionary mechanisms, and the creationists still appropriate and abuse it. If you publish anything about evolution, be sure to google your paper periodically — you may find that you’ve been unwittingly roped into endorsing creationism.
David LA, Alm EJ (2011) Rapid evolutionary innovation during an Archaean genetic expansion. Nature 469(7328):93-6.

The president has announced that he has an American birth certificate, like this was really an issue. If he thought this would end the yammering inanity, he was mistaken.
Donald Trump is preening.
He should have done it a long time ago. I am really honored to play such a big role in hopefully, hopefully getting rid of this issue.
Playing the role of a prancing moron who promoted the issue is nothing to be proud of, Donny.
Meanwhile, the Republicans are playing a game of pretending they didn’t never call Obama’s parentage into question, no how.
In a statement after Obama spoke, Republican National Committee Chairman Reince Priebus called the issue a distraction — and yet blamed Obama for playing campaign politics by addressing it.
“The president ought to spend his time getting serious about repairing our economy,” Priebus said. “Unfortunately his campaign politics and talk about birth certificates is distracting him from our number one priority — our economy.”
A Republican…chastising someone else for not being serious? Watcha gonna do, gomer, fix the economy by shutting down Planned Parenthood, praying, and giving rich people more tax breaks? Yeah, that’s serious.
I agree that Obama shouldn’t have wasted his time with this nonsense. Besides, you know that wingnuts are next going to argue that his birth certificate is a high-tech fake cobbled up by the geniuses at the NSA. Conspiracy theories simply do not die when presented with evidence — they absorb the evidence and claim that it’s further evidence of an even more elaborate conspiracy.

The existence of Ken Ham proves that creationism changes, therefore proving that it is not absolute, inerrant truth as he claims, therefore Ken Ham disproves creationism. QED. We can all go home now.

By the way, that’s not a bad diagram for illustrating the relationships of the various versions of creationism, except that the pedigree breaks down for Intelligent Design creationism, which is kind of a weird syncretic hodge-podge that accommodates both of the major branches of the creationist tree. It really ought to be shown as the warped, evil offspring of an unnatural act, an orgy of all of the critters in the bestiary. And then it also lies about its parentage (as would we all, if we sprang from such mindless chaos).

Oh, look what’s on the pharmacy shelves! It’s “medicine” for cute little babies! Everyone loves babies, and we want them to gurgle and coo and be happy, so when their widdle tummies make them cranky, we give them a little medicine to make them feel better.

Only…Brauer is marketing homeopathic “medicine” to kids — it’s not going to do anything. Those colicky babies are going to suffer and continue to cry and cry and cry, and the only change will be that Brauer will be a little richer.
Brauer profits off the pain of children, and offers nothing in return.
If you use advertising that exploits our susceptibility to sympathize with cute smiling babies, there ought to be some kind of backlash against them when it’s discovered that they are abusing those babies.

That open letter to the NCSE by Jerry Coyne really seems to have set the cat among the pigeons — it’s an amazing flurry of ruffled feathers. I don’t see how there’s any hope of reconciliation, either, as long as the apologists for religion continue to be as obtuse as they have been. Roger Stanyard of the BCSE is unloading furiously on Richard Dawkins right now:
We don’t entirely know Peter but this is very definetly an attack on both the BCSE and the NCSE by the “New Atheists” including Richard Dawkins. It’s very personal and nasty as well. Basically they want the NCSE and the BCSE to back “New Atheism” and many of the people signing the letter are very hostile to us including and working with people who are religious.
The highlighting is mine, but the stupidity is Stanyard’s. How often do we have to repeat ourselves? There is no goal of turning the NCSE or the BCSE into an atheist organization; we think having an organization that is honestly neutral on the religious issue is extremely useful in advancing the cause of good science education for all. We want the NCSE/BCSE to support neither atheism nor religion.
You know what? The atheists in this argument have a crystal-clear understanding of the difference between atheism and secularism, and are saying that the science education organizations should be secular. It’s these sloppy accommodationists who have allowed liberal christianity to become their default position who have violated the distinction.

Steve Novella is going to appear on the Dr Oz show today, and that promo has convinced me that Oz is an unforgivable charlatan. I’m swamped with work — we’re into the final two weeks of classes — so I don’t think I’ll be able to tune in, but any of you who do…report back! I’m sure Novella will acquit himself well, but it really looks like flaming woo central.

I have to agree with Ezra Klein’s explanation.
President Obama, if you look closely at his positions, is a moderate Republican from the early 1990s. And the Republican Party he’s facing has abandoned many of its best ideas in its effort to oppose him.

I read this story with mounting disbelief. Every paragraph had me increasingly aghast. It’s another case of physicists explaining biology badly.
It started dubiously enough. Paul Davies, cosmologist and generally clever fellow, was recruited to help cure cancer, despite, by his own admission, having “no prior knowledge of cancer”.
Two years ago, in a spectacularly enlightened move, the US National Cancer Institute (NCI) decided to enlist the help of physical scientists. The idea was to bring fresh insights from disciplines like physics to help tackle cancer in radical new ways.
Uh, OK…I can agree that fresh insights can sometimes stimulate novel approaches. Cancer is an extraordinarily complex process, but maybe, just maybe, the scientists studying it are so deep in the details that they’re missing some obvious alternative avenue that would be productive to study. I can think of examples; for instance, Judah Folkman’s realization that inhibiting angiogenesis, the process by which cancers recruit a blood supply from healthy tissue, would be a clever way to attack cancers beyond just bashing the cancer cells themselves. But then, Folkman wasn’t ignorant of cancer…he came up with that strategy from a deep understanding of how cancers work.
So I’m doubtful, but prepared to read something that might be new and interesting…and then I read Davies’ suggestion. Gah.
A century ago the German biologist Ernst Haekel pointed out that the stages of embryo development recapitulate the evolutionary history of the animal. Human embryos, for instance, develop, then lose, gills, webbed feet and rudimentary tails, reflecting their ancient aquatic life styles. The genes responsible for these features normally get silenced at a later stage of development, but sometimes the genetic control system malfunctions and babies get born with tails and other ancestral traits. Such anomalous features are called atavisms.
Charles Lineweaver of the Australian National University is, like me, a cosmologist and astrobiologist with a fascination for how cancer fits into the story of life on Earth. Together we developed the theory that cancer tumours are a type of atavism that appears in the adult form when something disrupts the silencing of ancestral genes. The reason that cancer deploys so many formidable survival traits in succession, is, we think, because the ancient genetic toolkit active in the earliest stages of embryogenesis gets switched back on, re-activating the Proterozoic developmental plan for building cell colonies. If you travelled in a time machine back one billion years, you would see many clumps of cells resembling modern cancer tumours.
The implications of our theory, if correct, are profound. Rather than cancers being rogue cells degenerating randomly into genetic chaos, they are better regarded as organised footsoldiers marching to the beat of an ancient drum, recapitulating a billion-year-old lifestyle. As cancer progresses in the body, so more and more of the ancestral core within the genetic toolkit is activated, replaying evolution’s story in reverse sequence. And each step confers a more malignant trait, making the oncologist’s job harder.
I’m almost speechless. I’m almost embarrassed enough for Davies that I don’t want to point out the profound stupidities in that whole line of argument. But then, there’s this vicious little part of my brain that perks up and wants to leap and rend and gnaw and shred. Maybe it’s an atavism.
Please, someone inform Davies that Haeckel was wrong. Recapitulation theory doesn’t work and embryos do not go through the evolutionary stages of their ancestors. We do not develop and then lose gills: we develop generalized branchial structures that subsequently differentiate and specialize. In fish, some of those arches differentiate into gills, but those same arches in us develop into the thyroid gland and miscellaneous cartilagenous and bony structures of the throat and ears.
It’s better to regard embryos as following von Baerian developmental trajectories, proceeding from an initially generalized state to a more refined and specialized state over time. Limbs don’t reflect our ancient aquatic ancestry in utero, instead, limbs develop as initially blobby protrusions and digits develop by later sculpting of the tissue.
Sure, there is an ancestral core of genes and processes deep in metazoan development. But Davies seems to think they’re lurking, silenced, waiting to be switched on and turn the cell into a prehistoric monster. This is not correct. Those ancient genes are active, operating in common developmental processes all over the place. You want to see Proterozoic cell colonies? Look in the bone marrow, at the hematopoietic pathways that produce masses of blood cells. The genes he’s talking about are those involved in mitosis and cell adhesion. They aren’t dinosaurs of the genome that get resurrected by genetic accidents. but the engines of cell proliferation that lose the governors that regulate their controlled expression, and go into runaway mode in cancers.
But even if their model were correct (which is such a silly way to start a paragraph; it’s like announcing, “If the Flintstones were an accurate portrayal of stone age life…”), it doesn’t help. We don’t have tools to manipulate atavisms. We don’t see any genetic circuits that can be called atavistic. The Flintstones might have made record players out of rocks, but that doesn’t imply that the music recording industry can get valuable insights from the show.
Oh, well, I shouldn’t be so negative. I’m alienating possible sources of work here. I understand the physicists have encountered some peculiar results lately. Have they considered bringing in a biologist consultant with no prior knowledge of particle physics? I have some interesting ideas that might explain their anomalies, based on my casual understanding of phlogiston theory and ætheric humours.

I do love me some Devo. Don’t neglect to check out the interactive video.
(Last edition of TET; Current totals: 12,262 entries with 1,350,652 comments.)
