I now have my first stack of papers to take home and grade for this semester. All joy is banished forever. That is all.
I now have my first stack of papers to take home and grade for this semester. All joy is banished forever. That is all.

I also liked “Thoughtleaders are more of a marketing gimmick than a philosophy,” and generally enjoyed this video immensely, in part because I detest the horseshoe theory and think that centrists are just polite fascists with a faint sense of shame.
I did disagree with one comment, though: “Islamists” are a thing. It’s a term used by Muslims and ex-Muslims to describe extremist religious fanaticism; it’s intent is to distinguish general, ordinary, non-fanatical Muslims from the raving loonies who use their religion to excuse violent, abhorrent behavior, so it is a useful word, just as “Christianist” is handy to distinguish, say, Theocrat Mike Pence from the more casual and benign faith of my mother.
A very nice summary:

Here’s a clip of Hurricane Irma shredding St Maarten:
Now imagine being on a boat in that weather.
Now imagine that that boat was little more than a barge built by a couple of guys with crude tools.
Now imagine that that barge is stuffed full of thousands of animals — it’s a makeshift zoo.
Now imagine that the weather is ten times, a hundred times, a thousand times more severe than Irma. OK, I can’t — I can’t even imagine being out in that disaster right now.
Now imagine that you’re such a gormless fool that you can believe that someone could ride out a storm like that in a floating barn.

Be safe, real people down there in the southeast.

Nature magazine has run a piece titled — brace yourself, it’s ridiculously bad — “Removing statues of historical figures risks whitewashing history”. It is subtitled “Science must acknowledge mistakes as it marks its past”, just to make it a little bit worse.
The objection is that people are clamoring to tear down a statue of a doctor and scientist, J. Marion Sims. How dare they question the honoring of a scientist?
The statues of explorer Christopher Columbus and gynaecologist J. Marion Sims stand at nearly opposite corners of New York City’s Central Park, but for how much longer? Both monuments have been dragged into a nationwide debate about memorials to historical figures who have questionable records on human rights. The arguments are long-standing, but were thrown onto the world’s front pages last month when protests against the removal of a statue of Confederate General Robert E. Lee in Charlottesville, Virginia, produced racially charged violence.
Last week, the Central Park Sims statue — one of many that stand in numerous US cities — was vandalized. The word ‘racist’ was spray-painted alongside his list of achievements, which include life-saving techniques he developed to help women recover from traumatic births. Yet many protest about the lionization of this ‘father of modern gynaecology’ because he performed his experiments on female slaves.
Yes, let us remember that Sims did save women’s lives. Sims pioneered a surgical treatment for vesico-vaginal fistulas (VVF), a common outcome of difficult labor that produced tears between the bladder and vagina and led to constant leakage of urine into the vagina. It was debilitating and shame-producing. Sims worked out a way to close off the fistulas. We don’t want to forget that!
Another thing we don’t want to forget is how it was worked out. That little line I highlighted up there, “because he performed his experiments on female slaves”, is minimizing what he did, and that also is a whitewashing of history, and failing to acknowledge a “mistake”, if we can call willful infliction of pain on unconsenting people a “mistake”. If we’re going to talk about the good that he accomplished, we also have to consider the evil of his method. I’ve read some justifications for his surgeries that say that because black slaves also suffered from VVF, it was legitimate that he experimented on them — they benefited too from his work! But let’s not forget that the reason he operated on these women is that he did not have to get their consent, and that part of his excuse is the belief that black people are less sensitive to pain.
And what he did was horrendous. Even acknowledging that all surgeries in the early 19th century were horrendous, he treated women like experimental animals. Here’s an account of his first experimental subject:
The enslaved women were not asked if they would agree to such an operation as they were totally without any claims to decision-making about their bodies or any other aspect of their lives. Sims used a total of seven enslaved women as experimental subjects; permission was obtained from their masters. They were in no way volunteers for Dr Sims’s research.
Nevertheless, Dr Sims was so positive that he was on the verge of making an astounding medical discovery that he invited local doctors to witness his first operation and what he thought would be a historical event. He performed his first operation on a slave-woman named Lucy.
Lucy was operated on without anaesthetics as Sims was unaware of the advances which had been made in this area of medicine. The surgery lasted for an hour and Lucy endured excruciating pain while positioned on her hands and knees. She must have felt extreme humiliation as twelve doctors observed the operation. Unfortunately, the operation failed as ‘two little openings in the line of union, across the vagina … remained although the larger fistula had been repaired’.
Lucy nearly lost her life, due to the experimental use by Sims of a sponge to drain the urine away from the bladder, as she became extremely ill with fever resulting from blood-poisoning. In recounting the episode in his autobiography, Sims says, ‘I thought she was going to die . . . it took Lucy two or three months to recover entirely from the effects of the operation’.
Sims’ method belongs in the history books, and no one is proposing erasing this protocol from the annals of medicine. But ignoring the suffering and degradation of the women in this experiment, as we have to do to think Sims deserves the honor of a prominent monument, erases a shameful era in our history, all while Nature protests that those who understand the full range of Sims’ actions are the ones doing the erasure.
It’s embarrassing, too, because whoever wrote this ought to know that the work of scientists isn’t honored with statuary. It’s honored with the work of those who follow afterwards.

I am a bad, foolish person, and now I’m feeling guilty. A few years ago, I made an appointment for a colonoscopy, and then what happened? Work happened, and I had to cancel my appointment. The prep work for the scan is unpleasant, and I knew it was going to mess up at least a day, and I couldn’t afford the time just then.
Worse, after canceling, I didn’t make another appointment. I’m just letting it slide. You know, that’s stupid. Especially after opening the latest issue of JAMA this morning and reading an editorial, Using Outreach to Improve Colorectal Cancer Screening. Early diagnosis of colon cancer makes a huge difference in prognosis, so you’d have to be an idiot to put it off.
Colorectal cancer (CRC) is the second leading cause of cancer death in the United States, with more than 50 000 deaths expected in 2017. Screening can reduce CRC mortality, and several methods of screening are available and recommended for average-risk adults aged 50 years to 75 years. Modeling studies suggest that several different methods of screening produce relatively similar levels of mortality reduction if there is good adherence to the underlying screening program.
Despite strong evidence of effectiveness, more than a third of age-eligible US adults are not up-to-date with CRC screening, with important disparities by ethnicity, income, education, and availability of a regular source of care. Currently, most CRC screening in the United States is achieved by colonoscopy. Studies of audiotaped encounters suggest that few clinicians and patients are having high-quality, shared discussions about screening options.
Increasing screening levels to greater than 80% has the potential to prevent an estimated 200 000 deaths in the United States in less than 20 years.
At least I’m not alone in my stupidity — a third of people in my cohort have been blowing it off. That’s no excuse, of course. Knowing that there are tens of millions of people in my situation does not make it less derpy.
So…I’m going to call in and make an appointment today. There’s a long wait time around here, so I probably won’t get in until Decemberish, which is all the more reason to call now.
Much of the editorial is about how health care providers can better inform and encourage people to get screened, and assessments of the effectiveness of various methods. I’m not a doctor, so I’ll just mention that I think outreach like Crispian Jago’s cancer diary helps me realize how important this is — and although it would be convenient to complain that my local hospital should have followed up with me after my canceled appointment, the truth is it’s all on me. And on you. If you’re over 50, contact your physician and make arrangements, if you haven’t already.
It’ll be fun! Weird liquid diets and spending a day purging your bowels so they can slide a camera up your butt? How can it not be exciting?

One side of the country is looking at Hurricane Irma rushing at them. I wouldn’t want to be in Florida right now.
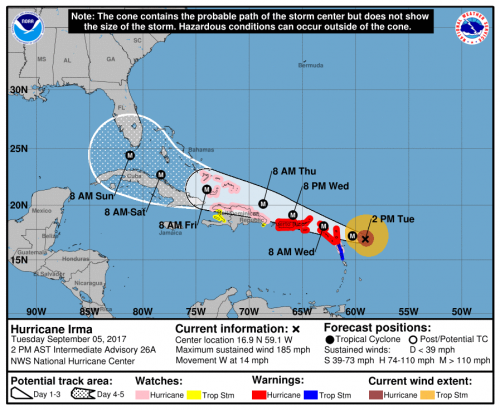
The other side of the country is on fire. I’m hearing stories from family members that it is snowing ash.
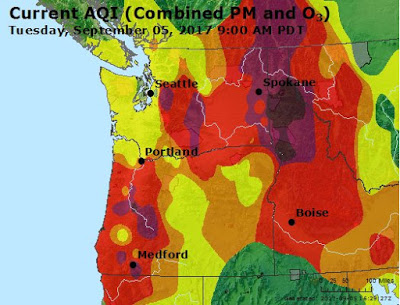
If only we could bring the two together for a day!
I’ll just mention that we’ve got clear skies and 20°C temperatures out here in the middle. Minnesota is perfect! Until December, anyway. Maybe November. Depends on your cold tolerance, I guess.

This is a guest post by Joshua Stein, a doctoral student at the University of Calgary and @thephilosotroll on Twitter.
There is a great deal to say about Molyneux’s The Art of the Argument, none of it good. The book begins with an introduction to logic that misnames (and misapplies) core concepts like validity, truth, and soundness (Loc 213), confusing how the “truth” relation is supposed to work (Loc 1572-1579), among other basic technical errors. I could dedicate an entire review to showcasing the bizarre and confused claims of the first quarter of the book; when I mentioned reading it on twitter, however, another graduate student reached out to me that he was going to be doing that review. Instead, I thought I’d do something a bit different (and, I hope, a bit more interesting).
In his opening moves, Molyneux differentiates between “Truth arguments” (Loc 68) and “Value arguments” (Loc 70). This roughly maps on to a philosophical distinction between descriptive and normative claims. “The window is open,” is a descriptive claim. “Close the window” is a normative claim. Some philosophers think that some descriptive claims may be normative, and vice versa. (For example, when my girlfriend gives me a look and says, “the window is open,” she is actually directing me to do something about it.)
In keeping with this bit of Molyneux’s analysis, I want to argue that the core argument of The Art of the Argument deserves both a descriptive and normative note (respectively): the argument fails and one who is committed to the argument ought to reject Molyneux’s views about libertarianism. Put another way, the argument is both wrong (a description) and those who accept it ought to reject Molyneux’s libertarianism. (Of course, I think everyone ought to reject libertarianism, but that requires responding to more serious political philosophers than Molyneux.)
Molyneux says we have an obligation to ostracize those who fail to meet certain philosophical standards (Loc 615-630). If I agreed on that point, I would encourage ostracizing Molyneux; it’s clear from an analysis of the first quarter of the book that he’s wildly philosophically incompetent. But I disagree with that tactic on both a pedagogical and moral basis; those who argue poorly can be taught to argue better, and we (as a community) are best served if those who can argue poorly are taught to argue better, so philosophy teachers ought to do that.
Perhaps the only reason many of us are aware of Molyneux is his insistence that various things are not arguments. This continues in the book. He notes “Taxation is not an argument, because you go to jail if you don’t pay.” (Loc 568) This is obvious, though it’s also obvious that dinner isn’t an argument, because potatoes aren’t premises. His positive account of what an argument is involves mixed metaphors and strange assertions. In his introduction, he insists that valid arguments with false premises are aren’t arguments, which is weird.
For the purpose of making this conventional, I just want to use a minimal working definition of argument. An argument is a series of premises adherent to some rules of inference that produces a conclusion. Molyneux often refers to “The Argument,” and so I will set aside that term for his definition.
The following is what I take to be a primary argument of Molyneux’s book, that “The Argument” is a primary condition for civilization.”
The Argument is the primary non-violent means of changing behaviors or attitudes. (Loc 530, 538)
1.1 A “non-violent” change in behavior or attitude is one where an agent changes their behavior without the threat of force.
Civilization requires principally non-violent means of changing behaviors or attitudes.
Therefore, the Argument is a primary condition for civilization.
One can see why (1) is compelling; it certainly happens in everyday life. If you and I disagree about whether we should go to the movies or the park, then I can make an argument in the hopes of influencing your behavior. Of course, there are lots of other ways one can non-violently compel behavior; Molyneux should know some of them, as their basic stock-and-trade of libertarian thought. For example, instead of presenting arguments, we can make an exchange, agreement, or promise that will lead to satisfying both our preferences.
With this in mind, it looks like (1) is just false on a strict understanding of “primary.” There are lots of other ways to modify behavior. Hobbes, Locke, and Rousseau all expound ways in which humans can develop contractual relationships by presenting their own interests and creating conditions agreeable to all signers. Rawls expands this to include even possible signers. This is a fairly straightforward view in modern political philosophy, and on this very broad sense of contractarian, it seems as though nearly all modern philosophers about it.
But Molyneux’s argument gets worse, because it turns out (2) is also false. Molyneux notes that “Freedom is freedom from force,” and maintains that civilization has to involve non-coercive engagement. The problem is, even if we take “non-violence” to be the absence of force, it is still hard to say what he means by “force.” After all, governments use force to address dissenters, and Molyneux hopes to avoid force by advancing a successful argument (Loc 1292).
The problem is that it is not clear at all when force is being exercised. Is “force” only the actual or possible commission of violence by another agent? Or can conditions “force” an individual to do something?
Suppose a cancer patient goes to see a doctor; the doctor has access to the chemotherapy drugs that the patient needs for treatment. Does the patient’s position force them to make a purchase? The patient isn’t in the same position I am when I look at buying a new pair of jeans; she can’t simply walk away. If she has access to a different doctor willing to provide the treatment at a lower cost, then there’s market possibility, but she’s dependent on the presence of alternatives and the good-faith pricing practices of sellers, precisely because she isn’t “free” to abstain. If she doesn’t buy the drugs, then she’ll die.
If contexts can be coercive, then plenty of instances of argument are coercive. Philosophy classes are coercive, though the stakes are a lot lower than relationship between a patient with an acute condition and doctor. Students who attend my ethics class require the course to graduate; they need to graduate to go into their chosen profession. Could they leave my class? Sure. But the quality of life for those students would suffer significantly, both in terms of career prospects and their own personal preference to pursue a particular sort of career. Is the mere contextual feature a matter of force? Sure. It isn’t violent, but it influences students to take some arguments more seriously than others; as a teacher, I have a responsibility to make that force work for the best interests of the student, but it is silly to deny that it is there.
Molyneux doesn’t address whether these instances count as force, but he does say some very confused things about force. He insistence that the primary mechanism of managing those who do not engage in civilized society is ostracism, but it’s not. It is force, whether forced imprisonment or some other severe consequence. Social contracts are enforced; social norms are enforced; an ostracism itself is a form of force. It isn’t as violent as hitting someone over the head with a club, but as an ex-Mormon about the impact of disfellowship on their life and it becomes clear that it is a powerful, and coercive, force.
As this line of analysis continues, Molyneux’s account looks even worse. Claims about the sensitivity of standards for argument to social factors, the role of concepts, and the like come back to bite him. But, up to this point, it’s clear that Molyneux’s argument fails, since both premises are false.
Molyneux’s argument fails (in part) because it can’t even meet his own basic standards of clarity of definitions. He writes, “Almost all human conflicts result from a lack of clarity in definitions.” (Loc 1173) As absurd as that evaluation of human conflicts is, the notion that definitions matter is right. Whether screwing up definitions of truth (Molyeux’s view should be, “Truth is a two-place relation between a proposition and the world.” Introductory level philosophy for objectivists about truth.)
The more interesting point in this discussion is that Molyneux’s argument is not just demonstrably false and a clear conceptual failure, but that if one takes the argumentative conventions seriously, they should reject the various libertarian claims he makes in the book. Some arguments are bad because they make rudimentary philosophical mistakes; that’s surely true. But Molyneux’s is especially bad because it intends to advance objectionable (and even laughable) theses that includes demonstrably false claims about interpersonal relationships and how they manifest justice in a society.
Much of the book rehearses a familiar strand of libertarian thinking; there are some arguments in the libertarian literature that are philosophically interesting, but Molyneux doesn’t touch on them.
Molyneux maintains that in a “free society,” i.e. one that conforms to the moral values he advocates, the success of individuals is responsive to the quality of their work. If Timmy works hard and Steve doesn’t, then Timmy will be more successful than Steve, but this argument trades on a few assumptions that are demonstrably false. These assumptions are that, in such a society, (1) individuals start off roughly on an equal playing field and (2) individuals’ improvement on the playing field is the result of hard work.
I want to focus on condition (2), because it’s failure is the most destructive to general libertarian arguments like Molyneux’s, but (1) is interesting. It is transparently false that a libertarian society entails that individuals will start in a roughly equal position. Over generations, in a state where individuals can accumulate wealth and pass it on within their family, some individuals are going to be positioned disparately in ways for which they are not morally responsible. If Timmy is born to parents with no educational background, low income and financial security, or other issues (e.g. alcoholism in the home; abuse; etc.) then he is in a demonstrably worse starting position than Steve, born to an upper middle class family, much less one of the Trump or Kennedy children.
Wealth inequality is a fact of any libertarian society, but also of most idealized liberal democratic societies; extremely disparate wealth inequality is only a feature of the former, which makes his own libertarian position untenable. Liberal democracy, at least in formulations offered by Rawls, aspires to putting Timmy and Steve and Baron Trump on a par (though not necessary an equal position), so that it’s plausible that their success is a result of hard work, rather than the circumstances of their birth.
The further problem, and the more egregious error, is that (2) doesn’t track either. A libertarian should be aware of this, but it is common to ignore. Suppose Timmy and Steve (starting on an equal playing field) go in to work for two different companies; Timmy works harder than Steve, the quality of his work is demonstrably better, but either due to errors of management or demand for the product, it goes out of business and Timmy is unemployed. Timmy is not at fault, but is now in a far worse position in his career than Steve, who has a secure position at a company that has invested in Steve’s jobs skills.
This is one style of case where someone’s economic circumstance is not responsive to hard work, but rather to external circumstances; we see this practically in contemporary coal mining, auto-manufacturing, and other fields that has seen enormous job loss through no fault of the workers. The workers then unjustly carry the economic burden of the company’s failure. This isn’t a bug that the libertarian position is equipped to result.
But Molyneux’s position gets even worse. Molyneux insists that a labor market responds to the quality of work, but anyone who has spent significant time watching folks get hired and fired knows that this isn’t true either. Labor markets are responsive to the traits that managers value. If Steve has some other trait that Timmy doesn’t (perhaps he’s a relative; or more sociable; or holds similar ideological views), and prospective employers value that feature, they may overlook the fact that Steve doesn’t work as hard or as well as Timmy. Circumstantial factors matter to hiring and firing.
If we hold the view that working hard and well ought to secure someone at least basic financial safety, a point Molyneux insists it will if the market is allowed to flourish, then the above demonstration shows (contra Molyneux) that a libertarian society will not, and cannot, address this.
A 45-year-old coal miner who loses his job when the mine closes down due to shifts to alternative energy, or an autoworker who loses his job to automation, isn’t faring worse than his friends because he didn’t work as hard. He’s faring worse because market forces he couldn’t have anticipated at 18 have devalued his skills. If libertarianism roughly entails not acting on those market forces and precludes redistribution of resources to compensate for the circumstance’s unfairness to those workers, then it does not provide an answer to this issue. After subjecting you, dear reader, to so much Molyneux, perhaps it is worthwhile to turn to a decent philosopher more attuned to social obligations to fairness and justice. In his seminal discussion, “Justice as Fairness,” Rawls gives a note sympathetic to much of Molyneux’s account of rational deliberation, but without the glaring philosophical errors of general reasoning and political philosophy outlined above. The result is an articulation of the basis of Rawls’s political liberalism:
“The fair terms of social cooperation are conceived as agreed to by those engaged in it, that is, by free and equal persons as citizens who are born into the society in which they lead their lives. But their agreement, like any other valid agreement, must be entered into under appropriate conditions. In particular, these conditions must situate free and equal persons fairly and must not allow some persons greater bargaining advantages than others. Further, threats of force and coercion, deception and fraud, and so on, must be excluded.” 1985. “Justice as Fairness” Philosophy and Public Affairs 14.3: 235
Note: Since I read this on Kindle, all of the citations refer to kindle locations in the book, rather than page numbers.

Hey, remember that Catholic guy who was writing a column about how atheists are atheists because of daddy issues? He’s back. He had to write a response because the intensity of atheist eye-rolling was annoying him.
In that post, I did not attempt to explain any proofs or evidence for the existence of God—that wasn’t the point. The point was to pose a question: Why would anyone hope against eternal happiness? Referencing a book by Paul Vitz, Faith of the Fatherless: The Psychology of Atheism, I posited one among many possible answers, namely that since one’s own experience with his father (or lack thereof) seems to have a tremendous sway in his perception of God, a bad father might lead a child to wish that God didn’t exist. That thought struck me as terribly sad. Thus, I made a very specific point that we Catholics should be compassionate toward atheists, hence the title. Frankly, I expected rebuttals to this statement along the lines of: “You’re wrong! I had a terrific father, and I’m an atheist! And the same goes for everyone in my college sociology class!”
I waited for this type of rebuttal to pour in. And I kept waiting. But my point went largely unaddressed.
But…but…that was exactly the point of my rebuttal! Many atheists have happy relationships with their fathers, and many Catholics have miserable relationships with theirs, and still remain within the faith. And also, if Jehovah is your role model for paternal parenting, you are seriously screwed up. But I guess he didn’t read my post, or it didn’t count, or something.
So now you might think his follow-up is to defend his position that atheists need pity because they have bad dads. You’d be wrong! I guess he decided his original proposition was indefensible, so he’s moved on to instead insisting that there is too evidence for his god, which is, ha ha, a so much more easily supported claim.
It’s a weird article, though, because he repeatedly refuses to actually defend that claim, but instead says he could have. It’s like a total non-defense. He’s hunkered down in a trench yelling at us, but ready to duck back down at an instant. It’s chickenshit Catholicism.
And his ‘evidence’ sucks.
I could have mentioned a very basic evidence: the existence of matter. How do you explain the existence of matter—stuff like diamonds, oxygen, and Saturn? Atheists frequently reply that if you put a monkey in front of a typewriter for an infinite number of years, he will eventually produce Macbeth. Their point is that given enough chances, a well-ordered universe will eventually just happen. But even conceding the conclusions of the “Infinite Monkey Theorem”, we are left with a prior question: Where did the monkey come from? The monkey might produce Macbeth, but nothingness doesn’t produce somethingness if you just give it enough tries. How do you explain somethingness? As many philosophers like Gottfried Leibniz, the inventor of calculus, have wondered: “Why is there something rather than nothing?” Leibniz concluded that this somethingness meant there must be a Creator.
We know how diamonds, oxygen, and Saturn came to be: reactions in the hearts of stars, and the condensation of gases in the formation of the solar system. The harder question is how did the universe start, what was there before the Big Bang, etc., and in all honesty I have to say that I don’t know. I don’t see how stating with certainty that an anthropomorphic being with intelligence and super powers did it is a reasonable answer, though.
My answer to “Why is there something rather than nothing?”, though, is to say that if there were nothing, there would be no one here to ask the question. That says nothing about what was the initial trigger for our existence. It leaves it as an open question, which I’ll trust the physicists to be able to someday answer, I hope, rather than the theologians.
I could have mentioned the existence of not only the material, but the immaterial. For instance, where in the world does conscience come from? Why does man have a notion of right and wrong, and why is that notion so similar across the historical and cultural spectrum?
Conscience, or the general idea of guilt, obligation, and empathy, is the product of activity in the brain. It’s partly built-in as a function of the theory of mind in social animals, and partly a product of social learning. We’d be lousy (errm, I mean lousier) at building communities if we lacked the ability to interact conscientiously. We have a notion of right and wrong that is constructed by our social environment in our plastic, responsive brains.
What else would it be? The product of angels?
I could have mentioned intelligent design.
You could have. I could have laughed harder.
Intelligent design does not help your case. It’s a circular argument. You’re job is to tell me what the evidence is that the universe is designed, and it does not provide new information to say that there is a hypothesis that the universe is designed.
I could have mentioned causality and the necessity of First Cause, or Uncaused Cause.
Yeah, yeah, right, while invoking an intelligent being who is an exception to that necessity, which means it isn’t really a necessity at all, now is it? I could also argue that the physical nature of our universe is also eternal, and therefore doesn’t require a first cause. Same answer, no hocus pocus with gods.
I could have mentioned plenty of evidence, but it would be rejected by convicted atheists. Atheists are certain God does not exist; that’s that. Of course, it’s impossible to prove that God does not exist, since you cannot prove a universal negative. True-believing unbelievers are, however, untroubled by that quandary. For many atheists—the verdict is pre-determined. Why bring evidence to a show trial?
Hang on there: in the very next paragraph, this guy then asserts that we are Catholics and it is our loving duty to spread the Good News of the Gospel.
He is certain that his god exists so he rejects all criticisms of his religion, while complaining that atheists are arguing out of prior conviction, and are asking him to provide evidence for his beliefs, which he can’t do.
I’m a hardcore atheist, but I’m not claiming to possess absolute truth, Catholics are. I’m saying that 1) believers have failed to provide an internally consistent definition of a god that does not contradict available evidence, and 2) have failed to provide specific evidence that their god is the best possible explanation. Saying that “things exist” is not evidence for “my specific model for why things exist”, especially when there are better alternative models that don’t have the shortcomings of the god hypothesis. There is a fundamental distinction there that they consistently fail to notice.
Oh, well. I look forward to John Clark’s future columns in which he ignores all the criticisms of the last one so he can flutter over to another non sequitur packed with irrelevant claims.

This is a wonderful 19th century photo of a famous person from the Pacific Northwest — the daughter of Chief Seattle, dubbed Princess Angeline. I knew about her when I was growing up, and Chief Seattle, too, since they’re such key figures in the history of the area, and I’ve seen this photo many times. That is a strong and dignified face.

But I’d never known how she came by such a European name, until now. It’s a genuinely cringeworthy story.
Born in 1820 in Lushootseed, near modern day Seattle, Kikisoblu (Kick-is-om-lo) was the first daughter of Chief Seattle, the leader of a Suquamish Tribe (Suquamish) and Dkhw’Duw’Absh (Duwamish). When American settlers arrived in Seattle, Chief Seattle befriended one of them, David Swinson “Doc” Maynard.
When the second wife of “Doc” Maynard, Catherine Maynard, saw the beautiful Kiksoblu, she said, “You are too good looking for a woman to carry around such a name as that, and I now christen you Angeline.”
Kikisoblu is a lovely name! In fact, all the Coast Salish place names that dot Western Washington are pretty and resonant — so it’s odd to see that kind of dismissal. And the Maynards have a reputation as being the early settlers who were most sympathetic to the natives. (Shhh, don’t tell anyone, but the pioneers that named a city after an Indian chief were mostly brutal, violent, and aggressive towards the people who lived there first — Seattle itself was a permanent collection of Coast Salish villages that had existed for about 4,000 years before the Europeans showed up. Surprising, I know.)

A child is brought to the US by their illegal immigrant parents; they grow up knowing nothing but America, go to American schools, have American friends, are fundamentally American. And then Donald Trump decides that, because of their parentage, they are going to be thrown out of the country and sent to a different country that they might not know anything about. He wants to repeal the Deferred Action for Childhood Arrivals program, which doesn’t go far enough in accommodating these kids. I think the DREAM Act, which would have allowed undocumented immigrants to earn citizenship with college attendance or military service, would be an excellent idea — we ought to recognize that there are people living here, who want to live here, who want to be part of this country, and that we ought to be welcoming them.
It is intolerably cruel to deny these young people anything short of full acceptance. They have committed no crime, yet Trump wants to make them suffer.
It is un-American to base people’s role in life on their ancestry — or at least, it is an overt rejection of the myth of American equality and opportunity. At least, the ideal that this was a place where you could advance yourself by studying and working hard, even if imaginary, was part of what we told ourselves made America great. The Republicans would rather your position in society was determined by inheritance, I guess.
There are a few moderate Republicans who see this, but far too many are as heartless as Trump. This anti-immigrant attitude that is sweeping through voters is chilling and horrible, too: our country is supporting an idea that the Nazis would have promoted as a matter of course.
These monsters must go. It’s not just an arbitrary political decision, it’s becoming a matter of defining the basic humanity of the American people…and the current regime is making all the wrong decisions.

