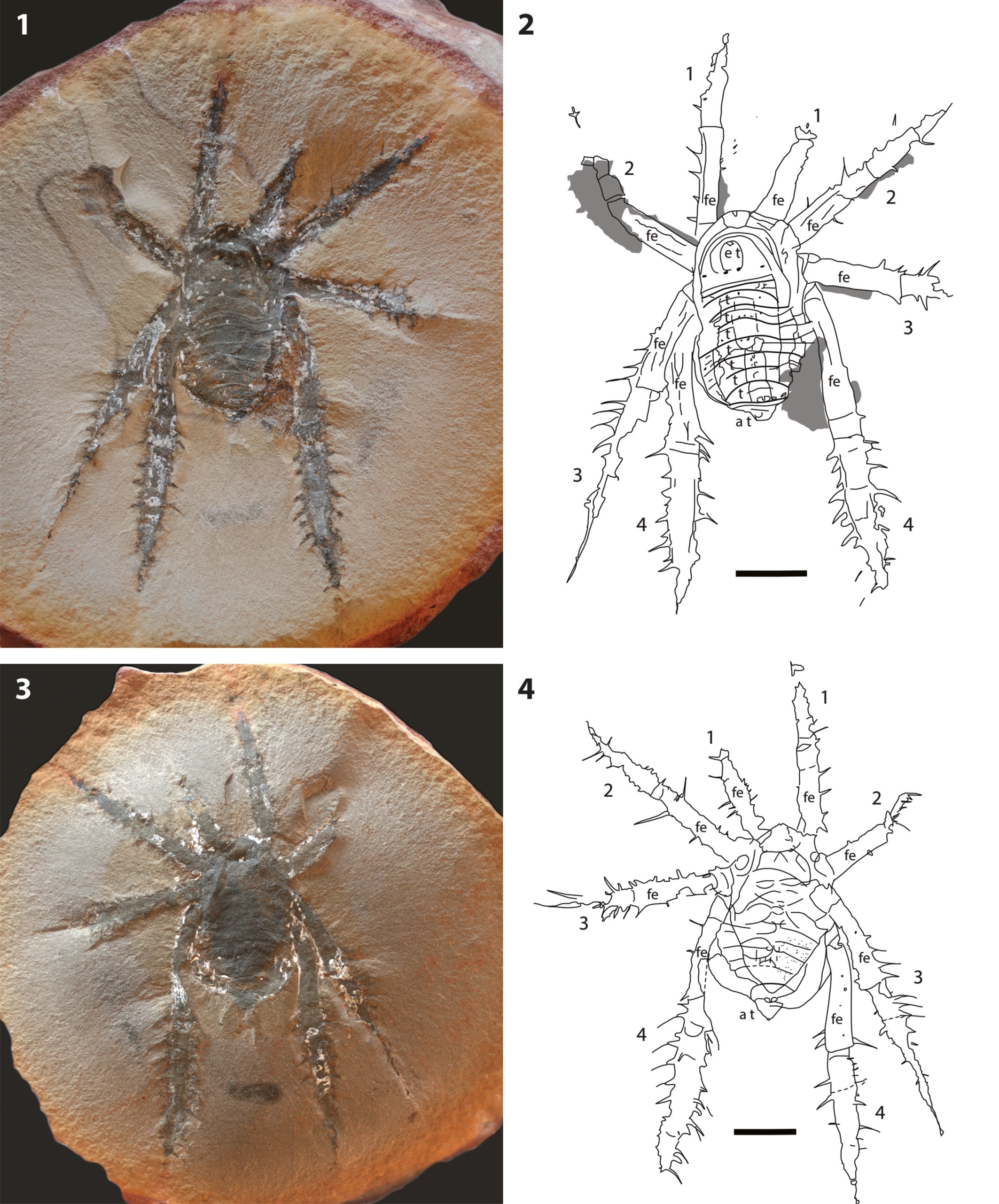
Our cat is a shameless coward that fears any kind of vermin, so useless as a mouse catcher. The spiders who live on and around my house are much more effective and relentless in taking out invaders.

Walking around the yard this afternoon I spotted four instances of bloody murder of pests. Good work!