My university offers courses in several languages with a relatively tiny population base, like Anishinaabe, the language of the indigenous peoples of the upper Midwest. I’m sometimes, but not often, asked why we bother, because these languages are so “impractical”, and it’s because they’re good for sharing the culture and history of groups of people, or because someone wants to understand their grandfather’s perspective (or the Teachings of the Seven Grandfathers). We should value the diverse ways of the multitude of different groups that have grown and changed throughout history. Unfortunately, many of these languages are dying out, and being replaced by a small number of “behemoth languages,” and we’re losing world languages at a steady rate — which is why it’s important to preserve as much as we can. This paper, The rise and fall of language diversity through the Holocene, tries to model the appearance and loss of languages over the last few millennia.
There are ~7500 languages signed and spoken in the world today, and they have a wide range of grammatical structures, vocabularies, speech sounds, signs, and usage rules. The study of language diversity has brought distinctive insights into how human behavior and cognition are shaped by ecological, social, and communicative pressures. The observable limits of linguistic diversity—features that are either universally present or absent in the world’s languages—have been at the core of what makes humans a distinctive species. Insights into language have also been used to understand other dimensions of human variation and homogeneity, including cultural and psychological traits. Over the past 500 years, all evidence points to a large systematic decrease in linguistic diversity, in parallel with the spread of a handful of behemoth languages, such as English, Spanish, Mandarin, Arabic, and Hindi. Nearly 5 billion people use at least 1 of the 10 most common languages, whereas half of the world’s languages are now endangered, and about four become dormant every year. This results in the loss of cultural and scientific knowledge embedded in minority languages and contributes to the unequal development in science, technology, medicine, and education that predominantly benefits speakers of the world’s largest languages.
I’m sure speakers of Anishinaabe recognize the definite advantages of speaking English, or maybe Mandarin in a few years, but they still want to retain a connection with their ancestors. Something similar happened to my immigrant great-great-grandparents — they spoke Norwegian, lived in communities of fellow Norwegian immigrants, and right up to my great-grandparents generation, were learning Norwegian and speaking it in church and family gatherings, but my generation has completely adopted the dominant English language of my region. We might still occasionally say “uff-da” or have lutefisk and lefse on holidays, but otherwise, we’re totally assimilated.
So about 4 languages a year are disappearing. The authors of that paper have attempted to quantitatively estimate the number of languages spoken over our history, and it’s surprising how much we’ve lost.
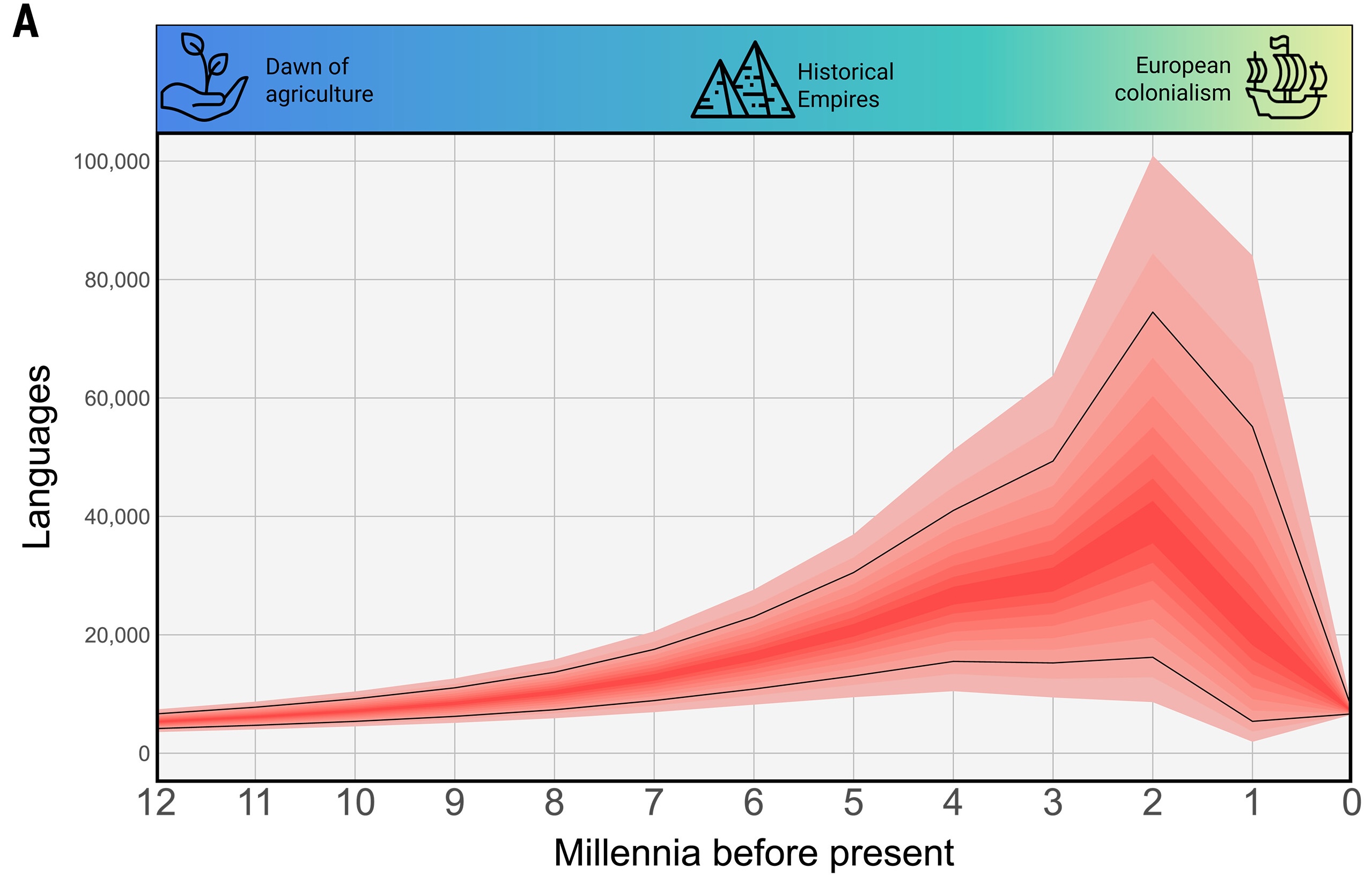
Fig. 4. Marginal results of our model aggregated across all conditions.
(A) The number of languages through the Holocene. Lighter shades of the curve correspond to progressively wider intervals comprising 10 to 90% of the predictions. Black lines indicate the interval containing 70% of the predictions.
Whoa. That’s a pretty sharp peak and a rather precipitous decline. The authors propose an explanation of what was going on.
Typically, the total number of languages has been increasing for most of the Holocene, mimicking the global expansion of human populations. This suggests that the rise of large-scale regional powers 4000 years ago did not generally and immediately deter the growth of linguistic diversity. Instead, most trajectories reached a maximum of 1000 to 3000 yr B.P. Although there is wide uncertainty in our estimates, most trajectories display an approximately 10-fold increase in the number of languages between the early Holocene and this point. This period, harboring tens of thousands of languages, has escaped previous theorizing.
In our model, this “golden age” was followed by an extremely rapid decrease in linguistic diversity, especially over the last two millennia. The process unfolded so quickly that the rate of language loss must have been even faster than the current pessimistic forecasts for the year 2100.
As populations grew, the number of languages also grew…until the rise of large empires and increased trade and communication, when a few successful dominant groups swallowed up the smaller populations. This model also makes the evolution of various languages non-adaptationist — some languages thrive, not because they are superior to other languages, but because they are piggy-backing on other properties of their population. Don’t bother trying to argue that English is better than Spanish, because what really matters is the strength and success of the population speaking the language. Languages don’t die because they’re inferior, but because they’re taken over by an expanding population.
It follows that this massive and biased reduction in linguistic diversity has shaped the relative abundance of different language traits. However, functional factors (such as learnability, efficient communication, or computational costs) are typically invoked instead to explain why some language traits are more common than others. But the intensity and recency of the linguistic diversity bottleneck make it a clear nonfunctional null hypothesis for understanding any skewed linguistic distribution.
Cool. I do have to envy linguists because at least they don’t get stupid demands like “Show me an Italian giving birth to a baby speaking Thai.” A lot of the elements of language evolution are similar to the properties of biological evolution, and seem to be equally incomprehensible to the ignorant.