This is weird. Perry Marshall has posted a complete transcript of our discussion on Unbelievable on his website. That’s actually useful, since most of us can read faster than someone else can talk. What’s weird is that he’s annotated it with his rebuttals, after the fact.
It’s like — and this has happened to me — you give a student an exam, and they do poorly on it, and they come in to your office to argue for more points not by pointing out errors in the grading (that happens, too), but by explaining to you how their understanding of the material was vastly superior to yours, and that you’d recognize that their answers were correct if only the professor had their deep understanding of cellular and genetic processes. In these situations, the student is always digging themselves into a deeper hole, and revealing that they don’t understand what they’re talking about at all.
For instance, he seems quite taken with Barbara McClintock’s work, and talked about it a fair bit. He got it all wrong. He claims that she was denying the existence of chance events in genetics — that everything was about patterned, engineered information, which is the damnedest interpretation of McClintock. He urges everyone to read her Nobel speech, which is good, but I’ve read her original papers and lecture on this subject in my genetics class. So it’s amusing to see him take a dig at me about it.
PZ doesn’t appear knowledgeable about McClintock’s work. Her findings perplex Darwinists because Darwinism has no grid for cells re-engineering themselves… but that’s exactly what they do in experiments.
McClintock was studying a phenomenon called chromosomal instability, in which the results of inheritance were not easily predictable. The opening sentences of her Nobel speech set the stage.
An experiment conducted in the mid-nineteen forties prepared me to expect unusual responses of a genome to challenges for which the genome is unprepared to meet in an orderly, programmed manner. In most known instances of this kind, the types of response were not predictable in advance of initial observations of them.
Note: unusual responses, a genome unprepared to meet in an orderly, programmed manner, responses that are not predictable. Most of us would read that and understand that it was going to be about chance events — events without predictable, programmatic, mechanistic responses. Not Perry Marshall! McClintock talks about stress-induced reorganization of the genome, and he leaps to the conclusion that she’s describing a teleological phenomenon in which the genome reshapes itself directly to address the circumstances, when every process she actually describes is about increasing variation.
For instance, he’s obsessed with transposition. He thinks this is engineering.
Transposition is when the cell moves a defined cassette of coding sequences and plugs it into a new location. [The “cassette” link shows transposition cassettes in experiments with zebrafish]
Transposition is not just inserting unspecified DNA sequences. Transpositions are by their very structure non-random, and would be even if there were no pattern to where insertions occur.
Aaargh. Yes. Transposition involves taking a chunk of DNA sequence and moving it to a different place in the genome. We can take advantage of that by making a gene of interest the ‘chunk’ and getting it to insert somewhere. Somewhere random. That’s the whole point. These are called jumping genes for a reason.
McClintock also described the bridge-breakage-fusion cycle. Marshall doesn’t understand it. He read the words in her speech and lacked the background to see what she’s talking about. Here’s the way it works.
You’ve got a chromosome with a duplication of a set of genes, as illustrated below. This allows for a misalignment of the homologous chromosomes, and for a crossover event to produce a peculiar chimera, a dicentric chromosome.
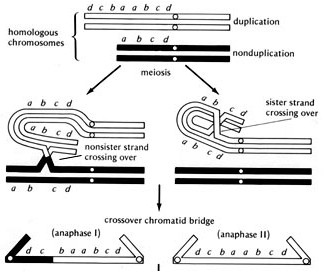
Notice that these chromosomes have two centromeres each. The centromere is a kind of handle that motor fibers grab onto and use to tow the chromosome to one side of the cell or the other during cell division. This chromosome has two, so what can happen is that one centromere gets pulled to the left, the other gets pulled to the right, and you see something called an anaphase bridge form between the two sides. It’s a tug-of-war, with the chromosome the rope stretched between two forming cells, and these opposite poles are pulling on it.
What do you think happens? The rope breaks. It breaks at some random point between the two centromeres.
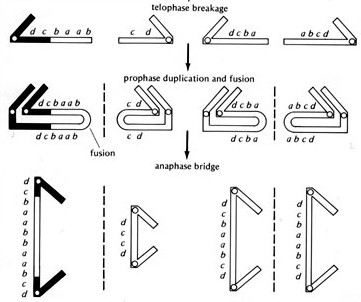
The breakage point is also typically a dangling bit of single-stranded DNA, with no telomeres. DNA repair mechanisms kick into gear and fuse the two dangling ragged strands of two homologous chromosomes back together, reforming the bridge so that when the cell divides again, it will break at a random point once more. And so the cycle begins again.
That’s why it’s called the bridge-breakage-fusion cycle. The chromosomes reform into a configuration that minimizes exposed, broken ends, but can’t last through cell division, so they’re constantly broken, reassembled, and broken again, producing a changing distribution of genes in each generation.

McClintock describes this in her Nobel speech. Marshall missed the relevant bits. She was trying to explain how a certain strain of corn developed variegations — that is, irregular, unpredictable patterns of streaks or patches in the tissue. It’s basically a random phenomenon, like the pattern of colors in kernels of Indian corn, and she predicted that it was caused by random breakages during cell division of a ring chromosome (a chromosome where the two ends are damaged, and are repaired to form a loop of DNA).
It was the behavior of this ring that proved to be significant. It revealed several basic phenomena. The following was noted: (I) In the majority of mitoses replication of the ring chromosome produced two chromatids that were completely free from each other and thus could separate without difficulty in the following anaphase. (2) Sister strand exchanges do occur between replicated or replicating chromatids, and the frequency of such events increases with increase in the size of the ring. These exchanges produce a double-size ring with two centromeres. (3) Mechanical rupture occurs in each of the two chromatid bridges formed at anaphase by passage of the two centromeres on the double-size ring to opposite poles of the mitotic spindle. (4) The location of a break can be at any one position along any one bridge. (5) The broken ends entering a telophase nucleus then fuse. (6) The size and content of each newly constructed ring depend on the position of the rupture that had occurred in each bridge.
What she’s saying is that crossing over between ring chromosomes can produce dicentric chromosomes, as in my diagram above, and that during cell division the bridge can mechanically rupture, and most importantly for this conversation, in point 4, “The location of a break can be at any one position along any one bridge”. That is, it can break randomly anywhere along the bridge, and then point 6, the gene “content of each newly constructed ring depend on the position of the rupture”. Each daughter cell gets a random selection of the genes along that bridge.
That’s the whole point. You’re trying to explain a random phenotype by looking for a randomization mechanism in the genome. McClintock’s triumph was being able to explain random variation within an organism that by convention ought to be genetically uniform by mechanistic processes like transposition and bridge-breakage-fusion.
As expected, Marshall has just dug himself a deeper hole with his rebuttal. I repeat: he doesn’t understand what McClintock was saying.

Hey PZ,
So I took college level intro bio maybe 8 years ago. I’m a doctoral student in Philosophy, so much of this debate is way over my head. My focus is on ethics and political theory, and I’ve just been a natural atheist for as long as I can remember. Anyway, I’m not getting the point, just for the sake of argument, why Perry being right ‘matters’ in any theistic/atheistic/ID sense.
“Most of us would read that and understand that it was going to be about chance events — events without predictable, programmatic, mechanistic responses. Not Perry Marshall! McClintock talks about stress-induced reorganization of the genome, and he leaps to the conclusion that she’s describing a teleological phenomenon in which the genome reshapes itself directly to address the circumstances”
Let’s just say the genome does reshape itself directly. So what? If I skin my knee, and address the wound, or if my dog skins her paw and addresses the wound, or a plant loses some leaves and addresses the damage, I don’t understand why that has anything to do with existence or non existence of a deity? I realize in the first two cases someone could say, ah but you’re intelligent, and then try to spuriously carry the knee analogy over into a ‘larger knee healer’, God. But I don’t understand why teleological processes at various levels of biological constitution, would grant theist anything. I mean if my immune system knows how to cure me without intelligence, then why can’t a cell have internal functioning processes to repair this genome damage? What am I missing? It always seems to me like animals, plants, and various organisms do have ‘teleological processes’ to a degree, even if there is no grander theistic goal behind them.
Big fan,
CB
Perry Marshall wrote:
oh, experiencing a few instances of errors over a very small sample set, producing no “practical” advantage is PROOF that evolution can not possibly result from stochastic events?
For all his software skills, i think he needs a little more education about statistics.
has he never heard of or even considered the question of “infinite monkeys randomly typing on infinite typewriters producing a work of Shakespeare”? Given how many cells and the number of trials, random errors is quite capable of producing extraordinary results and not just useless errors.
but what do I know, I’m just some random nerd who reads a lot and sometimes comments about the work of learned scholars as if I know better than people who work at it.
The idea that software science has to be ontologically tantamount to cellular science is a giant non sequitur, and metaphysically reckless.
anarchobyron: it doesn’t matter in the grand philosophical sense. Perry Marshall could come up with a video recording of a god poofing frogs into existence, and we’d just say, “OK, you’ve got data, we have to modify our ideas to incorporate that.” Similarly, if there were evidence for intelligent design, well, evidence and experiment trumps all, and we’d have to renounce our atheism and accept the facts. This isn’t about proving atheism.
It’s about conforming to reality.
The thing is that the evidence from McClintock’s experiments and observations does not support a purposeful and directed cellular response to stress. It says that cells repair damage as best as they can, but that they’re also basically thrashing about trying to fix random damage and producing variation, some of which will effectively deal with it, and others of which will not. There’s a probabilistic aspect that’s part of the story, and Marshall is denying that. So that’s why I correct him, not because randomness is some essential part of my worldview which must be defended at all costs, but because that’s what the evidence says.
And yes, there’s a kind of abstract teleology to cells — they have functions, and they live or die depending on how well they execute those functions. Cells reanneal broken ends of chromosomes because cells that don’t trigger cell death mechanisms that police the integrity of the genome. But there’s no plan to make a change that might improve the health of daughter cells.
As a quick aside, if I did see Frog’s appearing suddenly into existence, my first response [and my next dozen or so] would have nothing to do with a deity, let alone a judeo-christian deity. But that’s an aside. And I don’t really know what intelligent design evidence would even look like, since it always seemed to me to be a philosophical and metaphysical argument, incapable of scientific proof or disproof. Okay, now that that’s stated, and bracketed, I think Perry is wrong for other reasons.
Again, let’s just grant that he’s 100% correct in reading McClintock, you’re wrong, and in fact cells of repair in the way he says they repair. Again, so what? There’s no LOGICAL connection, that I can see, that says because cells repair this way, therefore intelligent design, or god exists, or god is the best explanation for that phenomena. Just as there’s no necessary connection between saying just because I repair my skinned knee in such and such a fashion, therefore god exists. Or just because Trees handle winter cold in such and such a fashion, therefore god.
I get that as a scientist you want the science to be understood correctly. That’s a fine and laudable goal. Don’t ever stop. But if you have to humor these ID wingnuts, it’s always easier, at least in the philosophical profession, to show why even if someone’s premises were true, they don’t get the conclusion they desire INSTEAD of quibbling over premises. Fine, let’s say McClintock did say what PM said she said. So what? ID hasn’t made a single step forward. Just shut the guy down on his own terms, then get back to the good science :)
Or am I missing something? (very probable).
Also, another aside. I see he says this:
“So if evolution is true then the universe is an open book and we can learn to our heart’s content.”
That’s quite obviously not the case, actually. If evolution is true, and there’s no reason to doubt it, the universe is a shut book, and we can learn to the degree that our biological constitution allows us to learn. In the same way a snake does not have access to parts of the universe we do, due to biological construct, we lack access to parts of the universe a different evolutionary chain could have taken an organism down.
Chomsky explains the point here at 2:33
Your problem is that you think I was making an argument that I wasn’t. I was not saying that McClintock’s story was this, therefore atheism.
I was saying McClintock’s story was this, therefore Perry Marshall is full of shit. It’s a much narrower issue.
As for this…
But she didn’t. He got her completely wrong. Why should I concede a falsehood to address a greater possible truth? A “truth” built brick by brick on lies is probably not. We should be concerned about epistemological validity at every step in our argument.
Oh no I don’t for a moment think you were saying McClintock X, ergo atheism. That’s just as silly as McClintock X, ergo ID/God. Like I said: “I get that as a scientist you want the science to be understood correctly”, and that’s great!
But it always strikes me that folks who do what PM is doing, are taking a belief, in ID and/or God, and then looking for areas of science where they can justify their belief. And so it just seems prudent to point out, look, even if he’s right about McClintock it doesn’t make his theism, or whatever he wants to call it, anymore true.
Hell, run both arguments at him. 1. You’re wrong about the science. 2. It wouldn’t matter if you were right about the science.
As a scientifically ignorant spectator, I listened to the debate, and read the transcript, and the whole time all I could think was “I lack the knowledge to ascertain whether PZ or PM are right on the science, but logically I can see it doesn’t matter one way or the other in terms of advancing ID, so PM is pulling a bait and switch or a red herring”.
I tried talking to this guy on the Unbelievable page and gave up. He is the textbook definition of ‘not even wrong’. I don’t have the time for it. Too much real work to do.
@y4dar #9
I also tried to engage Marshall, on his own web site, about his wacky software/biology analogies. His tendency to equivocate, evade, and ignore any relevant points gets tiresome quickly. At least he does not seem to just delete comments that he does not like.
By the way, I am a big fan of your work. Your explanation, on the AskABiologist site, of how ERVs are solid evidence for evolution, really clarified things for me. http://www.askabiologist.org.uk/answers/viewtopic.php?id=3914
@anarchobyron #8:
You’ve pretty much defined the purpose of the Discovery Institute. This has been known fairly unequivocally, though, for a couple of decades. They even submitted documents to court proving it!
I think part of the problem is the way we, as scientists, tend to talk about
cell behaviorthe phenomena we are describing. The language implies purpose even when we don’t intend it to. PZ did it in this transcript when he says “cells hate…” I did it myself in my first version of this comment by using the words cell behavior. Non-scientists associate words like hate and behavior with purposeful action.Creationists really ought to give up arguments from design. If there *was* a designer it was very clearly some combination of insane, incompetent, and/or intoxicated. Design arguments, at this point, *harm* the Abrahamic idea of a perfect God.
I really don’t think this is truly about science. Not at the heart of it. The creationists have a conclusion, pre-chosen, and they see science (and anything else they can get their grubby hands on…) as useful tools to pound their conclusion into other peoples’ heads.
This is a matter of philosophy, epistemology specifically. It feels like correcting them on the science is bolting the gate after the horse took off. These people are bullshitters, in the technical definition, which means they’re worse than liars because liars actually care about the truth value of their statements; bullshitters don’t, and will do and say whatever they feel is necessary to advance their cause.
and thus, you have entirely described Intelligent design, in function, theory and form.
nothing but a red herring.
I just listened to the show, and I don’t see why the Unbelievable people though he would make for a good guest. What he said mostly seemed like irrelevant rambling. A couple of points:
Boy, did he spend a lot of time trying to make a point that genetic errors aren’t provably random. I’m not sure what he was getting at with all this, and why he would think that it supports his Intelligent Design Creationism views. He went on and on about how you can’t prove that they’re random, I guess because there could always be some underlying cause that you haven’t yet discovered which would cause the behavior you’ve seen. But PZ, I wanted you to say that the behavior is consistent with their being random, and if there were an underlying cause, it would be up to him to show that, which he hasn’t done.
You mentioned once that he seemed to be thinking that a random process with a non-uniform distribution isn’t actually random – I had that same thought before you said it, and I’ve seen it a bunch with the mathematically illiterate.
His views of information are indefensible. I have an electrical engineering degree also, and the schooling I got taught me that information comes from randomness all the time. My own explanation for the layman is that information is simply a pattern that takes some time to describe. Something that takes longer to describe has more information. What is the shape of the coastline of the western US? That has a huge amount of information content, and almost everyone would agree that it was, or at least could be, created through natural processes.
He leans on that ‘randomness is unprovable’ thing hard in the comments there too. To the point it seems like the crux of his whole position.
It seems almost some sort of nascent Presuppositionalist style argument. “How can you say there’s nothing purposeful going on because its random? Randomness could be intentional! It could even be non random in a way we don’t understand yet and we’d never see it thanks to that conclusions. If you conclude some process is random, how do you know? Do you know all the outcomes across all space and time? Seems necessary to assume something is not random in order to describe any previously undiscovered process. And therefore everything is non random”.
…which is intriguing as a line of thought, I must admit, as far as twisting the use of ‘random’ goes. If totally bizarre at the same time.
I rather like both angles (“empirical” and “philosophical”) in the response to the similarly doubled claims, that seek to lead to a particular deistic invention and try to insert bunk into methodical, rational investigation. These two claims grope around for some place to grip, so I’m happy to see the ways in which they can be countered.
PZ is rightly uncomfortable with taking the rhetorical position that says “even if you were right, and you’re not, but if you were…”
That’s slightly coy and besides, why would someone concede facts that literally, physically can’t be conceded?
Oh, I’ll grant you the possibility that reality is a complete illusion — not that I think that, but if it was…”. It’s the way one speaks to someone who is halfway to understanding or just confused about some small aspect, not for someone who makes obfuscation their bread and butter.
Nevertheless, I also like the response that dares the claimant to deliver on his hocus-pocus as though it were true.
I’ve been investigating recently the ways in which bunk “Science” and “ideological” “Science” (for want of better terms) play parts in creating and reenforcing systems of control. Illegitimate control, I mean. They mimic religious hierarchies in some ways and the followers of such systems are as self-assured as any acolyte and equally without empirical or philosophical justification. Some prominent “atheist” writers come to mind.
I think that it is possible for a person to be completely ignorant of anything but their own bit of experience — no “Science” as such, no education (formal or otherwise), no ability to read or write even — and still respond to someone else’s claims about invisible forces and of those laundry lists supposedly handed down personally from the gods, always out of sight of an audience, conveniently. I think that the answer to claims about gods is ultimately personal: Do you want to subject yourself to someone else’s conceptions of (invisible) reality? So, one can say “but even if he was right, and he wasn’t, but if he was”, he’d be no closer, as anarchobyron stated, to supporting a case that puts his own gods in control of everything.
[blockquote]nothing but a red herring.[/blockquote]
I resemble that remark!