The Discovery Institute thinks axon guidance mechanisms are evidence for intelligent design. I think they just trawl the scientific literature for the words “complex” and “purpose” and get really excited about the imaginary interpretations in their head of papers they don’t really understand.
There’s no mention of evolution here, nor in the full paper in Science. The paper, however, does use a notable word: purpose. “These findings identify NELL2 as an axon guidance cue and establish Robo3 as a multifunctional regulator of pathfinding that simultaneously mediates NELL2 repulsion, inhibits Slit repulsion, and facilitates Netrin attraction to achieve a common guidance purpose.” In fact, they use it again in their concluding sentence:
Our results also show that Robo3.1 serves as an integrative hub: Its three diverse actions in response to three different cues — mediating NELL2 repulsion from the motor column, potentiating midline Netrin-1 attraction, and antagonizing midline Slit repulsion — act simultaneously, are mutually reinforcing, and serve the common purpose of steering commissural axons toward and across the midline. This multiplicity of mechanisms likely helps ensure high-fidelity steering of axons to their targets.
It’s one of those occasions in biology (not rare) when the term “intelligent design,” despite other merits, falls flat as a description. This is super-intelligent ultra-design.
Getting axons in the nervous system to their proper destinations actually is a very complex problem: much wires, many connections, wow. If you look at complex systems like the brain, you shouldn’t be surprised that the mechanisms are complex. And further, the functional requirements of those systems, which may require that Neuron A navigate to Target B in order for the pattern to work, it’s easy to say that the purpose of those mechanisms is to hook A up to B. It does not imply the existence of a designer, only the existence of functional constraints.
But also, they picked a system with which I’m fairly familiar. Way back in the 1990s, this is what I did: try to figure out the rules behind commissural neuron migration, the very stuff the DI is talking about. I was focusing on a cellular approach — I was observing neurons that grew across the midline to contact cells on the opposite side of the nervous system — and I reached some of the same general answers that more recent research has discovered. The question was why an axon would grow all the way across the nervous system to reach a target that had a closer equivalent right next door, on the same side.
Here’s a simple cartoon version of the problem. Neuron A is supposed to, has the function of, has the purpose of connecting to Neuron B; in the normal animal, it grows all the way across the midline to touch the contralateral (on the opposite side) B neuron.
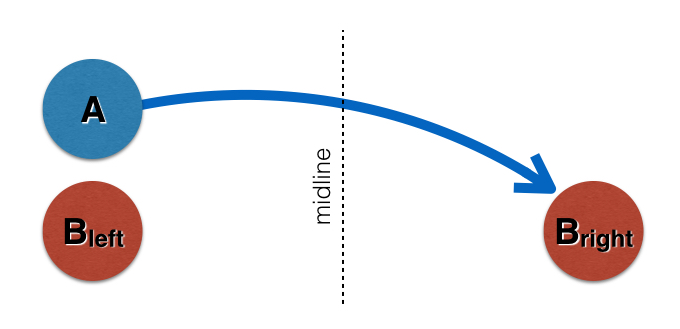
But the question remains: there’s a left B and a right B. Why doesn’t neuron A on the left side take the lazy shortcut and grow straight to the left B?

The answer we came up with in my work is that there is a hierarchy of interactions. That A finds the midline much more attractive than B at first, so it grows to the middle of the animal, and then, after a brief flirtation with the midline, changes its priorities to favor B cells after all, and just keeps growing across the midline to find the other B. (Actually, what we found that was most important in changing the left A’s affinities was contact with the right A, which arrived at the midline at about the same time.)
We worked that out with direct observations of neuron behavior, and also a series of experiments in which we killed various cells A would interact with. What we didn’t know at all was what molecules were involved.
And that’s where the Discovery Institute is so wrong. We had a cellular description, but when other laboratories in the late 1990s started discovering the molecular signals involved, molecules like Netrin and Robo and Slit, it was a wonderful revelation. It’s like how on one level, you can see a car and watch it run and figure out general things like wheels and steering, but when you get out the wrenches and start taking the engine apart, you can really see the mechanistic basis of its operation. Every step deeper into the guts of the problem tends to reinforce our understanding that it’s fully natural, and was built around natural processes.
The other big shift was that we could now generalize to other organisms and pick apart the evolutionary foundations to these mechanisms. I was looking at specific cells in the grasshopper embryo, and we could see that those very same cells are present in other arthropods, but we didn’t have the tools to do molecular comparisons. Identifying the molecules responsible meant that we could ask if they were present in other organisms, whether they were conserved, and whether these molecular processes were used in multiple cells, rather than just the few I studied.
If the Discovery Institute had looked just a little bit harder (or had not intentionally chosen to ignore all the papers that studied the evolution of axon guidance mechanisms), they might have noticed that there’s a very interesting literature on how these molecules evolved. There are plenty of papers that survey the evolutionary pattern of axon guidance mechanisms.
When did axons and their guidance mechanisms originate in animal evolution? Many axon guidance receptors (e.g. type II RPTPs, Eph RTKs, and the DCC, UNC5 and Robo families) are related to CAMs of the immunoglobulin superfamily, suggesting that axon guidance mechanisms evolved from signaling pathways involved in general cell–cell or cell–ECM adhesion in an ancestral animal. The simplest animals with nervous systems are cnidarians, which have isopolar neurons arranged in ‘nerve nets’; simpler animals (e.g. sponges, mesozoans) have no recognizable neurons. Thus, neurons and their guidance mechanisms must have evolved in a common ancestor of all metazoans, but after the divergence of sponges (Figure 1). Intriguing recent work suggests that sponges, which have no discernible nervous systems, nevertheless contain a diverse set of receptor tyrosine kinases and RPTPs [54,102]. Thus, many of these molecules could have evolved prior to (and may have been necessary for) the evolution of nervous systems in the urbilaterian.
Many axon guidance mechanisms are not only conserved at the molecular level, but also at the level of the body plan (reviewed in [103]). For example, netrins are secreted from ectodermal cells at the ventral midline of nematodes and insects and from the floorplate of the spinal cord of vertebrates (dorsal midline ectoderm, homologous to the ventral ectoderm of insects). Thus, in an ancestral animal, circumferential movements of axons or cells around the dorsoventral axis were probably oriented towards or away from a midline netrin source, and perhaps also from a midline Slit source. Studies in the coming years are likely to reveal the extent to which the patterning roles of other guidance mechanisms have been retained during the evolution of different body plans, and may help further outline the likely organization of the nervous system of our primitive ancestors.
These molecules are also multi-functional and play roles in other systems than the nervous system. They’re important in organogenesis and the maturation of the reproductive system, and are part of an interactive network of cell signaling molecules. It’s really complex, but what the DI doesn’t appreciate is that biology and evolutionary processes are really, really good at generating complexity. Look ot all the things the SLIT-ROBO system does!
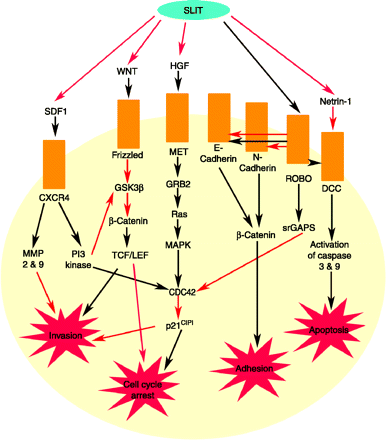
You might notice that they play a role in cancer signaling, too, but then everything does.
Once again, the Intelligent Design creationists completely miss the point. The work on these axon patterning systems has been deeply informed by evolutionary perspectives, while the DI is reduced to mining for mentions of “complexity” in papers, as if that somehow supports their ignorance-based position.
Chisholm A, Tessier-Lavigne M (1999) Conservation and divergence of axon guidance mechanisms. Curr Opin Neurobiol. 9(5):603-15. (Note that this paper came out very shortly after the discovery of netrins, by the fellow who discovered them — evolutionary biology has been part of this story from the very beginning.)
Dickinson RE1, Duncan WC (2010) The SLIT-ROBO pathway: a regulator of cell function with implications for the reproductive system. Reproduction 139(4):697-704.

Did you publish that work that you did in the early stone age? Just to get the timeline clear in the bibliography.
I’ve noticed something about the IDers and YECs; they tend to do very little research of their own, and primarily just trawl through actual scientists’ work and cherry pick whatever they think supports their position. Secular scientists who accept evolution are the ones who are actually doing all of the research into molecular biology and genetics that the creationists mine for out-of-context quotes. I’ve also noticed that their primary strategy is to simply look for words like “design”, or “plan”, and take that to mean a literal design or plan written by an intelligence. Human language is anthropomorphic, and it’s not unusual to describe non-human, unintelligent processes using human-like terms. For example, how often have chemists or physicists talked about atoms and molecules “wanting” to have stable electron configurations?
These kinds of posts almost make me wish I stuck with my life sciences fascination as a kid when I first discovered computers!
In fact, they use it again in their concluding sentence:
For a rather wide value of “concluding” (penultimate?) and/or “sentence” (paragraph?).
“Super” intelligent “ultra” design?
Very refute, much argue, wow.
Are we sure that was published by an adult?
I admit, I’m skeptical about the suggestion that there are people at the Disco Institute that know what an axon is.
It turns out to be extremely easy to disprove – even Einstein – by completely misunderstanding what they say. For example I can disprove Darwin by claiming that cell mutilation could NEVER turn a cat into a dog. And I’m right, too.
{NOTE: I had a philosophy professor in college say that exact thing!}
#1:
Growth cone dynamics during the migration of an identified commissural growth cone.
Myers PZ, Bastiani MJ.
J Neurosci. 1993 Jan;13(1):127-43.
Cell-cell interactions during the migration of an identified commissural growth cone in the embryonic grasshopper.
Myers PZ, Bastiani MJ.
J Neurosci. 1993 Jan;13(1):115-26.
The Discovery Institute never found any text or term that could not be twisted into whatever they wished for at the moment. After all, isn’t that what “discovery” means? To some anyway?
Yes, but is it RNA-mediated? Þ