All right, Larry Moran, why did you post about this paper now? I finished the unit on the origins of life in my cell biology class over a week ago, and this summary of the metabolism first model of abiogenesis would have been very helpful. I first gave them a review of redox reactions in chemistry, and then some general ideas about events in deep sea vents that generate a source of energy that early chemistry could have tapped, but this paper is full of specifics — probably a bit too heavy going for college sophomores, but they could have appreciated some of the diagrams.
Like this one:
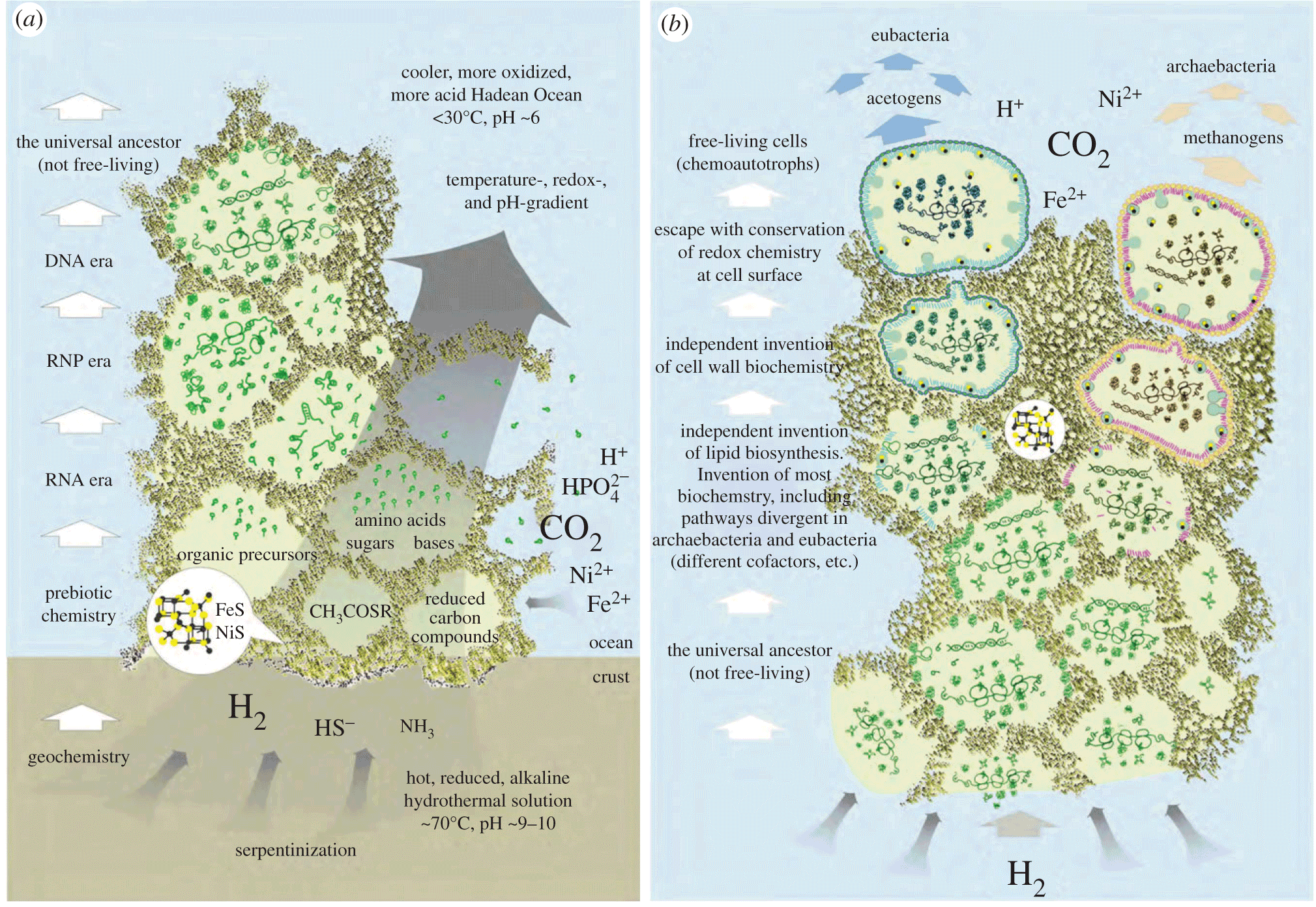
Also…the paper is open access! Everyone go read!
Sousa, F.L., Thiergart, T., Landan, G., Nelson-Sathi, S., Pereira, I. A., Allen, J.F., Lane, N. and Martin, W.F. (2013) Early bioenergetic evolution. Philosophical Transactions of the Royal Society B: Biological Sciences 368:20130088. [doi: 10.1098/rstb.2013.0088]

Just an FYI, your link to the paper has a problem.
Awesome. Being a physics nerd, and horrified by all the gooey bits inside that “life” stuff you weird bio’s talk about, I might not fully grasp it… but I look forward to being confused by it. :)
@chris61, 1
Link from the source:
http://dx.doi.org/10.1098/rstb.2013.0088
(I hope I’m not committing some horrible act by posting it in this way?)
Thanks Athywren.
With all due respect, I have to admit to doubting the paper, at least based on the diagram in the post. The image conflicts with a paper I have read about the evolution of bacteria, by Thomas Cavalier-Smith, which (among other things) argues, based on the invention of proteases and other transitions, that archaebacteria are clearly descended from posibacteria (the author’s term for gram-positive bacteria), with which they form the paraphyletic taxon, unibacteria (within which eukaryotes are rooted).
Also, Sousa et alia propose a “geochemically confined” ancestor, which sounds a lot (and in the diagram you reproduce in the post, looks a lot) like a proposal that Cavalier-Smith, to put it charitably, considers a contradiction in terms (under the headings ‘Lipid substitution in bacterial membrane evolution’ and ‘Cells, not genes, are the key units of selection and heredity’).
Not being a scientist, I can safely assume that you (and especially Myers) are more competent to judge, so is it just me or are Cavalier-Smith’s arguments and conclusions valid? And if not, why are they invalid?
And if Cavalier-Smith is right, how much of Sousa’s paper has to be wrong?
Edited to Add: I have noticed that Sousa et alia cite an earlier one of his papers (as reference 140), but they produce no actual evidence against a recent divergence of Archaebacteria, except to dispute two hypotheses about why it would have been advantageous.
As an aside, I read another of his papers, , which proposes that the first cells evolved from obcells, his term for ‘inside-out’ cells with the machinery attached to the outside surface and the inside used to store resources such as ions, pyrophosphate, et cetera.
Reposting to fix broken HTML
With all due respect, I have to admit to doubting the paper, at least based on the diagram in the post. The image conflicts with a paper I have read about the evolution of bacteria, Rooting the tree of life by transition analyses by Thomas Cavalier-Smith, which (among other things) argues, based on the invention of proteases and other transitions, that archaebacteria are clearly descended from posibacteria (the author’s term for gram-positive bacteria), with which they form the paraphyletic taxon, unibacteria (within which eukaryotes are rooted).
Also, Sousa et alia propose a “geochemically confined” ancestor, which sounds a lot (and in the diagram you reproduce in the post, looks a lot) like a proposal that Cavalier-Smith, to put it charitably, considers a contradiction in terms (under the headings ‘Lipid substitution in bacterial membrane evolution’ and ‘Cells, not genes, are the key units of selection and heredity’).
Not being a scientist, I can safely assume that you (and especially Myers) are more competent to judge, so is it just me or are Cavalier-Smith’s arguments and conclusions valid? And if not, why are they invalid?
And if Cavalier-Smith is right, how much of Sousa’s paper has to be wrong?
Edited to Add: I have noticed that Sousa et alia cite an earlier one of his papers (as reference 140), but they produce no actual evidence against a recent divergence of Archaebacteria, except to dispute two hypotheses about why it would have been advantageous.
As an aside, I read another of his papers, Obcells as proto-organisms: membrane heredity, lithophosphorylation, and the origins of the genetic code, the first cells, and photosynthesis., which proposes that the first cells evolved from obcells, his term for ‘inside-out’ cells with the machinery attached to the outside surface and the inside used to store resources such as ions, pyrophosphate, et cetera.
Pedanticspeaker, I am pretty sure that the proposal you attribute to Cavalier-Smith, that the archaea are not a separate clade from the bacteria, is a minority view.
The key pieces of evidence (simplified) that supports the general outline of Sousa’s model is the fact that the lipid membranes of archaea and bacteria and completely different, using different types of lipids with different synthesis pathways using unrelated proteins. The suggestion is therefore that the cell membranes seen in living cells are not all descended from a single ancestral cell membrane, but in fact was independently evolved twice. This in turn infers that LUCA might not have had a cell membrane, which evolved later, twice, in two separate lineages descended from LUCA. And if LUCA did not have a cell membrane, then it could not have been a free living organism, and would therefore need to be geochemically confined in some way.
Pertaining to this statement: “Cells, not genes, are the key units of selection and heredity”
An argument certainly can be made that this is true today.
However, when considering abiogenesis, we are considering, among other things, the origin of the first cell. Cells cannot be the key unit of selection and heredity until the first cell appears.
How did the first cell appear? Is a cell a simple enough thing that it can spontaneously arise? If not, then some mechanism must bootstrap simplicity to greater complexity in order to produce the first cell. Evolution (ie natural selection with heredity) is one such mechanism. And if so, then before the first cell, the unit of selection and heredity must be something other than a cell – ie the various components that would later combine to become the first cell.
To say that the cell “being the key unit of selection and heredity” must mean that selection and heredity applying to things simpler than cells is thus impossible and cannot be considered as a mechanism for producing the first cell is the same intellectual error as saying because “all cells come from other cells” a-la Pasteur means that the first cell could not have arisen abiotically and therefore must have been created.
I don’t pretend to understand most of either the paper referred to in the OP or the paper linked by pedantic speaker but it seems to me that the former is arguing for an origin of life based on conditions on earth prior to life and extrapolating forward while the latter is arguing by extrapolating backwards from today. Their conclusions are based, it would appear, on somewhat different sets of assumptions.
Athywren@2: there’s no problem linking to a paper, even if it’s not open access (as this one is). You could be violating copyright if you email someone the paper itself (or a large chunk of it), but linking to the source is fine. It is up to the provider to determine how to police access to the content.
pedanticspeaker, that first Cavalier-Smith article doesn’t really address abiogenesis — it’s a proteinomic analysis of existing organisms used to reconstruct a family tree of living creatures going back as far as the technique allows…to well after the evolution of cells with nucleic acid libraries. I don’t know enough about protein transition analysis to judge the paper technically, but it does seem to be a sound principle. I’ve only read the abstract to that second Cavalier-Smith paper, but I have to say that I think its second sentence is flat-out wrong (“Only selection at a higher level than for individual selfish genes could power the cooperative macromolecular coevolution required for evolving the genetic code.”) and is both a straw man (even the most ardent selfish-geneist would reject the idea that evolution only works on individual genes and never on functional groups of them) and is an Argument from Personal Incredulity which neglects the wealth of research showing that sophisticated co-operation can evolve among non-co-operative entities.
But while that sentence irritates me, the core of Cavalier-Smith’s argument is about the chronological development of proto-life structures (i.e. membranes, codons, etc.) whereas the Souza paper is about prebiotic chemistry and is more focussed on thermodynamics and substrate availability. I don’t think it matters all that much if there are some contradictions between the two papers given they are dealing with quite different aspects of a process that occurred billions of years ago under very different conditions to what we can observe today and left very few “hard” traces of what happened. Especially as Cavalier-Smith has a long track record of changing his tree-of-life diagrams as new evidence comes out (this meant as praise, btw, not a criticism) — so I very much doubt Cavalier-Smith would be too concerned about a new paper contradicting some of his earlier ideas…he’ll just take the new evidence and hammer out an updated clade diagram.
Extrapolating backwards from existing life gets us to LUCA, but really can’t get us further back than that. You have to extrapolate forward from initial conditions to get to LUCA. But you need to know what LUCA is likely like to have a target to extrapolate towards, which is what extrapolating backwards gives you.
In the end we need both processes.
Disagreement between individual researcher’s interpretation of the results of the different methods is what we would expect in a field that is on an active frontier of research.
It is virtually guaranteed that they will both be wrong in at least some aspects. With luck, there will be bits and pieces that are correct and help us move the research paradigm forward.
Amphiox@7:
Yes, but the problem with abiogenesis is that this logical conundrum exists for whatever we decide is the fundamental unit of selection. If you think cells are the fundamental unit, as Cavalier-Smith does, then the obvious point of attack on the problem of abiogenesis is on how cells first came into existence. If you believe molecular replicators are the fundamental unit of selection (as I do), then the problem is working out how molecules become self-replicators. That is, whatever the relative merits of cell-selection vs. molecule-selection vs. whatever-selection, all of us have the same hurdle to leap with regards to abiogenesis.
It looks like a good paper, and I’ll definitely be reading it, but hasn’t that diagram been around since at least Martin and Russell’s 2003 paper “On the origins of cells: a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells” (not open access exactly, but it does seem the Royal Society has made the full text freely available; link on the right hand side)? Koonin and Martin’s 2005 paper “On the origin of genomes and cells within inorganic compartments” in Trends in Genetics has the same diagram consolidated into a single whole. That paper is not open access, either, but a Google Scholar search will reveal a downloadable PDF. :)
As a high school biology teacher (in Thailand, so I don’t have to bother with any creationist nonsense trying to invade my classroom), I have been using those diagrams when teaching abiogenesis for several years now. I’m a big fan of the Alkaline Hydrothermal Vent hypothesis, as it so neatly solves the problems of a long term energy source (which you get from pH and thermal gradients), raw materials (which you get from the serpentinization of the hydrothermal vent fluids as they circulate through the crust), and concentration (provided by the compartments in the rock and the convection flow due to the thermal gradient). I am far from an expert, but so far this hypothesis seems more complete and coherent than other models I’ve read about. :)
With respect to amphiox at comments 6 and 7:
Firstly, are you suggesting that membranes made of “completely different” lipids cannot be descended from a the same ancestral membrane? If I may quote the first paper I mentioned,
Secondly, did you really think that I was unaware that isoprenoids are made by different enzymes than fatty acids? Cavalier-Smith actually argues that it “weak[ly]” favours a derived nature for archaebacteria, because:
Thirdly, in context, Cavalier-Smith was arguing that the last common ancestor was a cell (with two distinct membranes, within or beside chlorobacteria, et cetera); certainly the last common ancestor of archaebacteria and eubacteria had an essentially complete genetic code (considering how similar they are in all known lifeforms) and several hundred separate proteins at least (he cites a source that claims 683 protein fold superfamilies are shared by eubacteria and archaebacteria). It was clearly a complex organism. This is the particular proposal he was discussing, so I do not know if it applies to all “geochemical confinement” proposals, but:
Regarding chrislawson at comments 10 and 12:
Firstly, I had assumed that it would matter whether biological methanogenesis evolved before or after the first cells.
Secondly (regarding “I have to say that I think its second sentence is flat-out wrong”), I think that his argument in the second paper is that there needs to be some sort of structure to keep the genes together; otherwise, they would be irretrievably dispersed. In the paper, he outlines how he thinks obcells could turn into the earliest cells, so I assume that he means very late protocellular life if that helps clarify things.
Also, I just saw Alex Samaras’s comment at #13, mentioning the exact same paper on “inorganic compartments”. Cavalier-Smith does not seem to impugn the paper’s relevance to and validity regarding the simple self-replicating molecule stage of abiogenesis, but if he is right, they are not to be mistaken for proto-organisms:
I admit to being partial to Cavalier-Smith’s own favoured hypothesis, but I admit there are others that are far more exciting: For instance, if Cairns-Smith is right about silicate minerals as the earliest replicators, then they might still exist.
As a reply to pedanticspeaker, and keeping in mind I have not yet read the paper described in this blog post…
I don’t think any of the authors of those papers are calling the compartments proto-organisms in the sense of individual compartments being like individual cells. What they are arguing is that such rock near thermal vents acted as sort of biochemical reactors building starting from basing inorganic molecules and building up into larger and larger biomolecules eventually leading to an RNA macromolecule replicator. Not in individual compartments but throughout the rock. Once the first replicator formed, it would have had plenty of raw material to work with and so would start replicating more and more throughout the rock, with imperfect replication leading to variation, and so selection. This is the origin of heredity and the origin of biological natural selection (with the first traits under some kind of selection probably being fecundity, fidelity, and longevity). This is the RNA world. Over time, as perhaps raw ribonucleosides get used up faster than the vent system can make them, there might be selection favoring replicators that happen to be with ribozymes that can synthesize more nucleosides, such that replicators which replicate themselves and the ribozymes that help make more nucleosides get selected for. Or, perhaps in another case, replicators paired with ribozymes that break down certain RNAs for raw materials, and selection in that case favors replicators that also replicate those ribozymes.
Then stuff happens, genetic translation evolves, and RNA world becomes RNP world with proteins doing the work and RNA moving away from being ribozymes to being protein coding genes. The invention of reverse transcriptase leads to the use of DNA as information storage, and we arrive at LUCA. LUCA is not thought (in those papers) to be a single cell or a single compartment in the rocks, but rather a system of DNA based genetic replicators with a common genetic code and the main tools for DNA replication, transcription, and translation already in place, not to mention genes for some of the metabolic enzymes spread across such rocky compartments. LUCA is not a thing, it is a system. It is a population.
The proposal in these papers regarding the origin of cellular life is that LUCA precedes the first cells, and metabolism for making phospholipids had not been invented yet. When it was invented, it was invented in two separate, independent ways. And one possible reason for their invention was for ATPases and ion pumps to take advantage of chemiosmosis. PLOS recently had a really interesting paper regarding this hypothesis: A Bioenergetic Basis for Membrane Divergence in Archaea and Bacteria.
With the invention of phospholipid metabolism and membrane construction, this system would eventually evolve into the two separate branches of cellular life.
Now, I’m not saying that this is what happened. My point is, at no point do the authors argue that the individual compartments should be regarded as individual “cells”. The importance of the compartments is that they serve to concentrate the molecules coming from the vent long enough to react and accumulate larger and larger molecules over time. This is a big deal. The concentration problem is one of the main problems with many hypotheses regarding abiogenesis (does Cavalier-Smith address it? I haven’t read any of his papers in awhile, and the last ones I did read several years ago had not addressed it). The other main problems are an energy source, a continual source of raw materials, a way to get your life-like stuff into lipid membranes, and the system with all of those things being stable enough over a long time for life to have a chance to form. The hydrothermal vent hypothesis neatly addresses all of these things. Now, that doesn’t make it “true”. But, as a hypothesis, it fits the requirements of the things which need explained most quite well.
In my humble opinion, of course, and as best as I’ve understood the papers I’ve read. And, of course, I am not a real biologist, just a very interested amateur. :)
With respect to Alex Samaras at comment 16:
I really suggest you read Obcells as proto-organisms: membrane heredity, lithophosphorylation, and the origins of the genetic code, the first cells, and photosynthesis.
I do not think that he argues against “inorganic compartments” as a solution to the concentration problem, just that they are not sufficient to attain the complexity of cells, let alone the LUCA/cenancestor (he reckons that it was a photosynthetic bacterium with ~2000 genes and a double membrane, and considering that archaebacteria and eubacteria share over six hundred protein fold superfamilies, their common ancestor probably had well over a thousand genes), and that attempting to do so requires very strange logic. His own hypothesis for the origin of RNA replicators is in fact also based on hydrothermal activity, albeit in different circumstances:
Further, he argues that it would likely have been a polar or tundra climate, where prebiotic membranes and early RNA molecules would be more stable and “periodic freezing, as well as drying, could concentrate precursors without limit.”
In terms of the other main problems:
He hypothesises that the energy source was pyrophosphate generated by the effects of intense heat (eg, by volcanic action) on apatite or other phosphate minerals but he admittedly assumes a supply of RNA precursors.
He addresses the lipid membrane problem heavily in the paper, which is about how a proto-organism could have come into existence and evolved to the point where it became the first cell: In essence, he starts with
There are plausible reasons to add an amino acid tail, for instance to attach itself to phosphate mineral surfaces and hence concentrate it even further (but this would be “weak in salty environments”), and a longer hydrophobic tail would allow it to attach itself to abiotically synthesised membrane vesicles, and he argues that that would have been advantageous:
Also, where do we draw the line between populations and organisms? Chromosomes? Obcells? True cells?
Cavalier-Smith argues that there would be selective advantage in forming a chromosome well before the first true cell, and with as few as six genes (because “[a]s Maynard Smith and Szathmáry (1995) cogently argue, even as few as six cooperating replicators would probably be sensitive to such parasitism by selfish variants.”), so even assuming he is in error by a factor of ten and there is no selective advantage until more than sixty genes, the first chromosome would almost certainly predate the LUCA/cenancestor.
I’m reading the paper right now. So far it’s awesome! Best part yet:
At least some of Cavalier-Smith’s arguments are addressed in the paper.
I’ve read it now.
The paper is largely a review paper that presents a few new ideas in a lot of context. That’s why it has 246 references! As far as I can tell, the exact proposed metabolism is new, while the alkaline-vent hypothesis as such is not.
Uh, that’s what open access would mean. But no, I’m sitting at home and the full text is not free. (Apparently the price depends on the country the reader is in.)
Like everything else in phylogenetics, it’s a parsimony argument: there are so many things (Sousa et al. listed several) that bacteria and archaea do in fundamentally different ways that it’s easiest to assume that LUCA lacked these things.
Sousa et al. didn’t mention this; point for Cavalier-Smith.
Sousa et al. instead mentioned an idea for how the double membrane could have come about.
That paper, not cited by Sousa et al., does not contain the number 683 anywhere; the occurrences of 68 are limited to a bootstrap value and to pages in references, and 83 doesn’t occur at all, so if 683 is due to typos, you’ve made at least two. The last sentence of the abstract is instead: “During the course of the study, it emerged that there is a core set of ≈50 folds that is found in all 174 genomes and a single fold diagnostic of all Archaea.”
Sousa et al. found 106 genes to be homologous across Archaea and Bacteria (though some were lost a few times). Relevant paragraph:
“The list depicts what sorts of functions might have been present as genetically encoded functions in the common ancestor, allowing generously for loss and keeping in mind that we cannot readily tell how much LGT has contributed to those gene distributions. With those caveats, the list tends to reflect a last common ancestor that had ribosomes, the genetic code, bits and pieces of cofactor biosynthesis, bits and pieces of amino acid biosynthesis, bits and pieces of nucleotide biosynthesis, and a fully fledged ATPase that could convert an ion gradient into chemically accessible high-energy bonds. The paucity of obvious components that would generate an ion gradient is striking. The list in [T]able 2 is remarkably consistent with the view that the genes listed arose in an environment (an alkaline hydrothermal vent), where geochemically produced proton gradients existed and only had to be tapped. The presence of several tRNA modifying enzymes in the list is also congruent with our arguments about modified bases.”
Numbers in parentheses mine:
(1) Yes. For Sousa et al., that’s not a bug, but a feature. (2) Sousa et al. briefly described an idea for how division might have worked anyway; in the process, this idea describes how the double membrane of most bacteria may have come about. This definitely needs more work, though. (3) Correct. The unit of selection is the selfish gene which needs to survive and replicate in an environment that consists in no small part of other selfish genes. (4) The claim is mistaken, and Sousa et al. didn’t repeat it. However, it doesn’t matter anyway; full compartmentalization is not a requirement for selection and an increased diversity of complexity.
I don’t understand why you didn’t read the paper first. ~:-|
Several years ago it was suggested that DNA came long after LUCA – and appeared in viruses first… As the paper you cite mentions, the DNA replication machinery seems not to be homologous between Bacteria and Archaea.
Nice! :-) That paper, from August 2014, cites the one by Sousa et al. (June 2013), and greatly expands on one of the many things it said, presenting them much more clearly. :-)
Link doesn’t work.
Seriously?
It has long been thought (Nature or Science, late 1990s…?) that the gene cassette for photosynthesis has been laterally transferred several times. That’s how the heliobacteria got it, for example.
With respect to David Marjanović at comment 19:
The 683 number comes from this figure, with the caption,
According to the diagram, Archaea share 654 protein fold superfamilies with both eukaryotes and eubacteria, and a further 29 with eubacteria alone; only forty-nine were found in all of the genomes studied, but the sample includes “many [bacteria] which have reduced genomes”. And I never said it was cited by Sousa, I said it was cited by Cavalier-Smith.
Here. And if that fails, I also linked it in comment 4/5. I double-posted to fix HTML errors, but the link works in both posts.
According to Cavalier-Smith, it does not need to be invoked at all. Back in the first paper I referenced in comment 4/5, he wrote
Also, I looked for the very vague citation you gave, and I did not find much of relevance. I mean, I did learn about viral mutualism and that there are now five known carbon fixation pathways, but I do not think I found the paper you were looking for.
David Marjanović @# 19:
pedanticspeaker @#20:
Google scholar (and the references for open-access and otherwise available reviews of phtosynthesis genetics) suggests that the first discussion of horizontal gene transfer of photosynthesis genes was in Nagashima 1993, Nagashima 1997, and Igarashi 2001.
The article published in Science might have been Raymond 2002 (PDF), whose abstract reads: