On Wednesday, I’d had a little private pity party, moaning to myself how I really disliked my teaching schedule and would never do this again. You see, this semester, thanks to a sudden schedule change, I was teaching two very different classes back to back — I’d finish lecturing in genetics, and immediately have to swivel and scuttle off to teach introductory biology. I like to have a little break between my classes, a time to reorient myself, review the material, put my feet up, sip a little wine (OK, I don’t do the last bit, but I can dream.) I have been spoiled.
Thursday I was a guest at the local high school. Yikes. One class after the other, all day long. You get your lesson plan all mapped out well ahead of time, because once that first class launches at 8:25, you are on a fixed trajectory all day long, with only a few minutes between classes. Forget moments of reflection, don’t even think about the imaginary glass of wine, because a succession of students are going to march in and occupy your classroom.
I don’t think I could cope with teaching high school. Much respect to those who do — you are all overworked and underpaid.
On the positive side, though, it was a pleasant experience…for a day. Just a day at a time. The big difference between college and high school is that college students are generally so damned serious. They’re paying out big bucks and accumulating a substantial debt to be here, and classes are their job, while professors are the bosses armed with the scourges of exams. At the 7th and 10th grade classes, I started talking about spiders, hands were raised, students would spontaneously offer wild accounts of their spider experiences, they’d ask question after question, it would sometimes get a bit raucous. Their enthusiasm was wonderful.
Now how to get the college students that fired up…I think I’ll have to kill all the exams. Abolishing tuition would help, too. I’ll get right on that for next year.
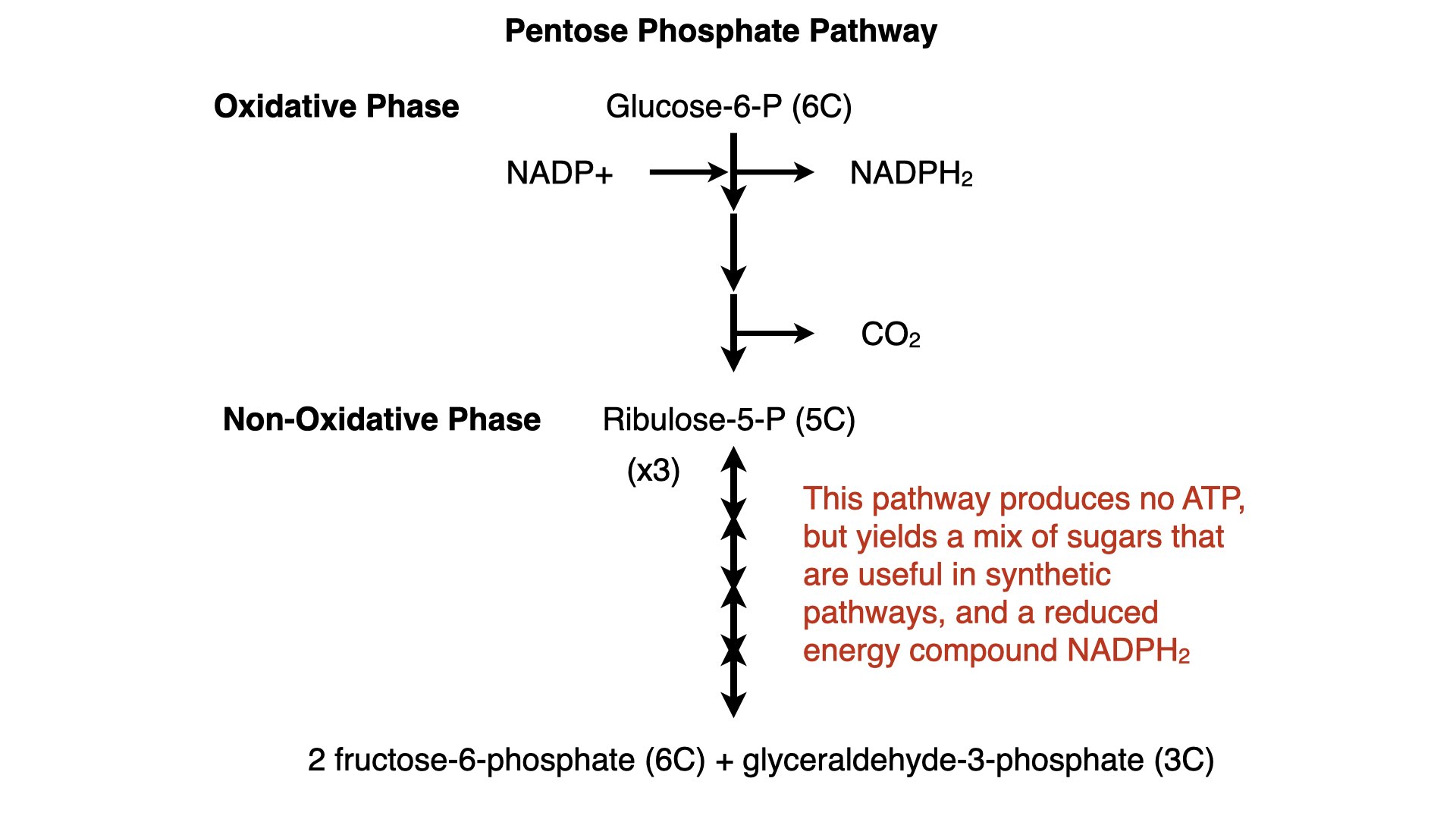
By the way, I also got to peruse their textbooks, briefly. There’s been a change there: they weren’t using Miller & Levine anymore! They’d switch to something called Inspire Biology, which looked fine, but different: lots of short, choppy segments with exercises to make the students think, less of a narrative, more for short attention spans. That isn’t bad, I could see how you could use textbooks like that to customize how you teach.
They did still use the familiar Miller & Levine lab manual and praised it highly.
For those who don’t know, Miller & Levine’s Biology was, for many years, the ubiquitous text I’d see in every high school student’s backpack. It was kind of like Campbell Biology at the college level. I’m seeing a lot more textbook diversity in the last decade or two as publishers seem to have realized that owning the rights to a popular textbook is a cash cow. For them, not the authors.