I love it when creationists decide to attack evolution by way of developmental biology. Yes, come into my parlor, said the spider to the fly. Our home is our terrible weakness, I’m sure you’ll be able to frolic and thrive in our little domain…heh heh heh <twirls mustache>
This fellow, David W. Swift, whose knowledge of development is maybe an inch deep and obtained entirely from information taken from basic textbooks, followed up by a conscious misreading of a few scientific papers with an eye to extracting only bits and pieces that support his conclusion, has decided that embryology is evidence against evolution.
It is quite well known that at a relatively early stage the embryos of a wide range of vertebrates look similar – a so-called ‘phylotypic stage’ – and this is generally regarded as evidence of common ancestry.
What is not well known is that before this stage the early embryonic development of vertebrates is very diverse – right from the earliest stages – which clearly refutes their common ancestry.
He’s partly right. It is well known that there are embryonic similarites between vertebrates, and of course that’s evidence for common ancestry. There is a lot of evidence for common ancestry, and that is one small piece of it.
He’s wrong when he says that it is not well known
that early development is more diverse. Any developmental biology textbook that discusses comparative embryology is going to tell you that. We developmental biologists ALL know that. It’s been a hot button topic of discussion for at least 50 years, probably longer. Mr Swift perused a select bunch of papers and just skipped over any that brought the topic up, so he could pretend he was the perceptive creationist who noticed, for the first time, a ‘weakness’ in evolutionary theory? Such arrogance. It’s especially annoying because he says nothing novel that he couldn’t have found prominently discussed in the scientific literature.
Also, the known evidence that early development is a complex process with diverse patterns of execution in different organism does not refute common ancestry. The creationists seem unable to get into their heads that a short-sighted process driven by chance as a core mechanism, running over millions and billions of years, is going to generate complexity and diversity by its very nature. Observing that something is complex and diverse is evidence that an evolutionary process created it, rather than an engineer.
That doesn’t stop him from expanding that claim into a whole tedious paper, published in the Discovery Institute’s fake journal, Bio-Complexity. The only difference between it and the short blurb on his website is the wordiness.
It is well known that the embryonic development of vertebrates from different classes (e.g., fish, reptiles, mammals) pass through a “phylotypic stage” when they look similar, and this apparent homology is widely seen as evidence of their common ancestry. However, despite their morphological similarities, and contrary to evolutionary expectations, the phylotypic stages of different vertebrate classes arise in radically diverse ways. This diversity clearly counters the superficial appearance of homology of the phylotypic stage, and the plain inference is that vertebrates have not evolved from a common vertebrate ancestor. The diversity extends through all stages of early development—including cleavage and formation of the blastula, gastrulation, neurulation, and formation of the gut and extraembryonic membranes. This paper focuses on gastrulation, during which the germ layers originate and the vertebrate body-plan begins to form. Despite its key role in embryonic development, gastrulation occurs in fundamentally different ways in different classes of vertebrates. The inference against common ancestry becomes progressively stronger as more is discovered about the genetic and molecular mechanisms that implement development. It is increasingly evident that these are of such complexity that it is unrealistic to think that undirected variations (random mutations) could produce constructive changes to development, such as those required to account for a diversification of development from that of a common ancestor, especially while retaining a similar phylotypic stage.
He’s focusing on gastrulation, which certainly is an interesting phenomenon. Gastrulation is a process by which animals form three embryonic layers from two, and there are multiple ways animals do it: by involution or a kind of folding/migration, or by delamination, or cells leaving a layer to reconstitute another one, and it’s also strongly affected by maternal investment. Animals that pack a lot of yolky goodies into their eggs have different patterns of cell division and movement than animals that produce small eggs with little yolk, so animal life history and ecology plays a significant role.
But that’s nothing new. It’s not a significant obstacle to comprehending evolution as the origin of the diversity (again, evolution is really, really good at coming up with diverse solutions, and also complicated Rube-Goldbergian strategies). Somehow, all these really smart scientists have been looking at this problem for decades, and they haven’t been leaving the evolutionary camp in droves.
For example, Bill Ballard was writing about exactly these issues for a long, long time. He was talking about it when I was a grad student and earlier. He has all kinds of papers specifically discussing the diversity of gastrulation in vertebrates, and he was particularly peeved at people who assumed that embryological homology can be traced all the way back to the earliest stages of development. These were vestiges of an “antique homological theory”, he said, that is, Haeckel’s recapitulation theory, and everyone needed to get over it.
It may be that the cells of representatives different classes of chordates or different phyla are behaving in such different microenvironments and being controlled by such different genes and forces that the analysis of their morphogenetic movements will be as difficult to compress into a single account as the description of their early embryonic stages has proved to be. It seems wise, in the meantime, to avoid assumptions of uniformity drawn not from precise observation but from antique homological theory. We should be cautious with the use of terms that have become more and more loaded with implied meaning while they were becoming less and less definable.
He doesn’t reject evolution because of this complexity, though. He suggests that “it may be” that cells in different organisms may be “controlled by such different genes and forces that the analysis of their morphogenetic movements will be as difficult”. Maybe there isn’t a clean, simple, single principle driving gastrulation, but a whole welter of divergent processes that will be difficult to sort out. That is not contrary to evolution at all!
Also, he wrote that in 1976. That was before evo-devo appeared on the stage, and maybe there will be some comprehensible unified principles establishing a unity in gastrulation, once we start combining embryology and molecules (hint, hint).
David Swift, writing in 2022, has no such excuse. In fact, he’s going to cite several well-known evo-devo scientists while completely ignoring their explanations. The bulk of his paper consists of pulling out drawings of the gastrula stage of various organisms, and saying, “See? They look different, therefore evolution is false.” He’s so focused on superficial comparisons with no deeper understanding that he shoots himself in the foot without even noticing. Pay attention to the last sentence in his diatribe.
In view of their morphological similarities, it is understandable that the phylotypic stages of different classes of vertebrates were interpreted as homologous and as evidence of common ancestry. However, this apparent homology is refuted by more detailed embryological evidence; despite their similarities, the phylotypic stages are formed embryonically in profoundly different ways. The straightforward conclusion to draw from this radical diversity of their early embryonic development is that it shows the vertebrates have not evolved from a common vertebrate ancestor. This conclusion can be avoided only if there are credible explanations for how such diversity of early development might have arisen from the development prevailing in a common ancestor (whether or not similar to present-day cephalochordates) in an evolutionary way, via changes that (i) had a realistic probability of occurring, (ii) maintained viability, and (iii) offered, in most cases, significant advantage that could be favored by natural selection. Further, to be taken seriously, such explanations can no longer be based solely on putative morphological changes, but must take account of what we now know about the genetic and molecular mechanisms through which embryonic development is implemented.
Holy crap, YES. We have to look at the genetic and molecular mechanisms. So why doesn’t Swift talk about them at all? He has one throw-away image that he doesn’t discuss, and he quotes important scientists like Rudy Raff, but he doesn’t seem to understand what they say at all — or more maliciously, edits out anything they say contrary to his perspective.
For instance, he ends the paper by citing Raff.
It is more than 20 years since Raff wrote: “One might reasonably expect mechanisms of early development to be especially resistant to modification because all subsequent development derives from early processes”, and the more we find out about how embryonic development is implemented at the genetic and molecular levels, the more it reinforces this commonsense conclusion. Many other authors have also commented on why we would expect early embryonic development to be resistant to change (for examples see Irie and Kuratani). Yet, when it comes to the diverse embryonic development of presumed homologous organs or body-plans, the usual assumption is that their early development must somehow have derived from that of a common ancestor, no matter how improbable the changes required, rather than accept the plain inference that the similar organs etc. are not homologous, at least not in an evolutionary sense. This expectation seems to reflect an ideological commitment to the theory of evolution rather than an objective assessment of the embryological facts.
You know by now that when a creationist partially quotes something from a real biology paper, it’s worthwhile to look to see what they intentionally left out. Here’s that Raff paper.
One might reasonably expect mechanisms of early development to be especially resistant to modification because all subsequent development derives from early processes. Traditionally, features of early development and conserved larval stages, even between phyla, have been regarded as strong homologous characters for the inference of phylogeny. The division of animals into protostome and deuterostome superphyla is based on the ideas that embryonic similarities are homologous and have been largely immutable over hundreds of millions of years (Raff 1996). A view of development from an evolutionary perspective is both more confounding and more interesting. Early development is highly evolvable, even among closely related species. The evolutionary portrait of ontogeny may be that of an hourglass, as shown in Fig. 1. In this diagram, embryos of two related species follow different early developmental trajectories, but converge on a similar phylotypic stage. It is important to note that the phylotypic stages of related organisms bear major features in common, but also have evolved significantly (Richardson et a1 1998). Divergence in post-phylotypic developmental trajectories yields variant adult species morphologies, as suggested by von Baer. The divergence of pre-phylotypic stage pathways can be extreme. For example, polyembryonic parasitic wasps have a bizarre early developmental pathway that does not resemble typical insect early development. Polyembryonic development produces 2000 embryos from one egg. Nonetheless, these secondary embryos develop via a characteristic insect phylotypic stage (Grbic et a1 1998)
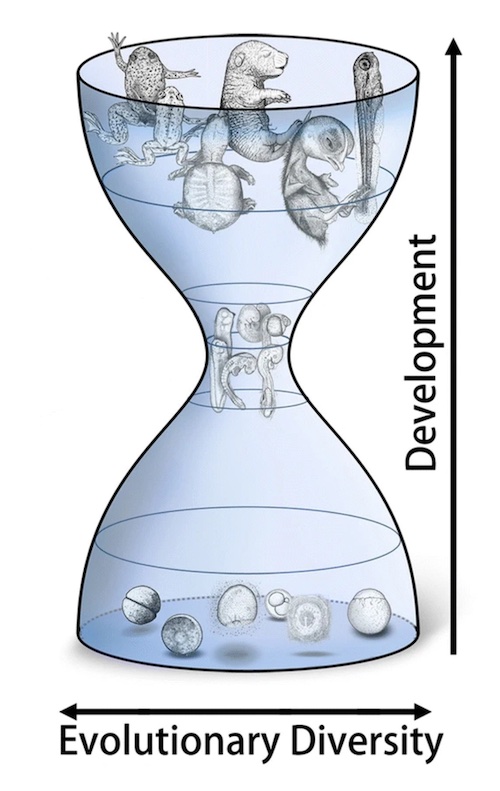
I guess he only read the first sentence, because the rest of the paragraph is pointing out the value of the evolutionary perspective. He also mentions the developmental hourglass, which Swift includes in his paper, which Swift fails to point out is not a creationist revelation, but a routine illustration of the phenomenon of early divergence which every goddamn developmental biologist knows about.
Raff also goes on to explain why this does not refute evolution.
With respect to Van Valen’s view that homologues represent a continuity of information, we find that some of the processes that underlie the evolution of development can confound what we mean by ‘continuity’ as well. Should all this be a cause for despair? No. Biologists have historically used homologies to trace evolutionary histories and phylogenetic relationships. A deeper understanding reveals that this cannot be a tidy programme. That is on the face of it unfortunate, and has generated much hand wringing on the usefulness of the homology concept. However, where developmental homologies are difficult to identify because of process shifts in ontogeny, we are actually being told interesting things about evolution. Ambiguities in development of homologues in embryos reveal where and how evolutionary changes occur and thus, although confounding, are difficult Rosetta Stones needed to understand how evolutionary novelties arise
That’s the same point Ballard was making! The complications are interesting, they are informative about the evolution of the process, we shouldn’t expect a tidy, simple program of development.
Furthermore, one of the cool ideas that emerged from evo-devo is the observation that hey, different organisms are using the same molecules in early developmental processes, like gastrulation. Swift makes the error of talking about the epithelial-to-mesenchyme (EMT) transition, a common process in which cells leave their tidy sheet (epithelium) to migrate as loosely aggregated mesenchyme to new destinations. It occurs in all kinds of developmental processes, as well as in cancer. Here’s all he says about it.
Behind the above changes that occur at the cellular level are
of course the genetic and other molecular mechanisms that effect these changes. Progress has been made towards elucidating these, and some are depicted in Figure 18.
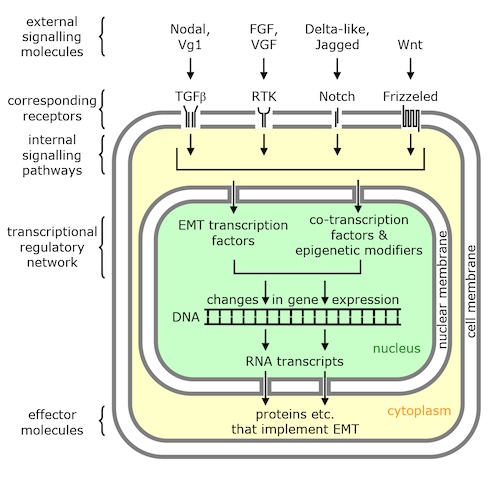
This is a strange self-own. He finally mentions molecules, and shows an image that lists a bunch of molecules, and I looked at it and was confused. Not because I didn’t understand it — those are all molecules I know very well — but because the diagram was ambiguous. What is this? Human cancer tissue, or a fruit fly embryo? Because it applies equally to all of them. Wnt/Frizzled (he misspelled it!), Delta/Notch, etc.…these are old friends. You can find them if flies and zebrafish and people. Read the Debnath paper Swift took it from, and as it goes through the molecules, it’s repeatedly saying things like “Snail was first identified in Drosophila melanogaster,” “Twist1 was first identified in drosophila,” “ortholog of human GATA6 acts as inducer of EMT in Drosophila endoderm,” etc., you mean Swift didn’t notice that we have molecular homologs of all these genes central to gastrulation that are present in multiple phyla? Curious blindspot he’s got there.
If you are seriously and honestly reading the developmental biology literature, as Swift is not, you can’t avoid this. It’s everywhere. Developmental biologists are taking these processes apart using the tools of molecular biology, and practically every paper acknowledges the way every organism is using the same molecules (and also ackowledging differences!) to carry out what may look like different cellular events. Nobody is shy about pointing this out, so it takes a kind of willful ignorance to overlook it. Here, for example, I pulled out a paper I’ve been reading about polarized shapes in epithelia. Notice the conclusion.
The recent demonstration that a similar, or even the same, Fz/PCP signaling pathway regulates both polarized cell shape changes in flies and CE in vertebrates suggests that this is an evolutionarily conserved mechanism. Strikingly, the conservation extends beyond the core Fz/Dsh signaling module in PCP and includes vertebrate homologs of other genes involved in Drosophila PCP [40–43]. Thus, although the end result of cellular polarization is distinct in different tissues and organisms, the underlying signaling pathways and mechanisms seem to be highly conserved. First, specific aspects of Fz/PCP signaling and their interaction with other PCP genes are likely to be similar between flies and vertebrates. Second, specific Fz/PCP-regulated processes such as activation of Rho, Rho kinase, myosin and JNK seem to be features that are shared between Drosophila and vertebrates. Taken together, therefore, there is growing evidence for the conserved function of Fz/PCP signaling and the conserved involvement of other primary genes in both epithelial PCP in flies and vertebrates, and in CE in vertebrates.
Swift is ridiculously stupid, lacking any real knowledge of the subject he’s writing about, yet it still manages to get published in an intelligent design creationism “journal”. They have no standards at all.
Ballard WW (1976) Problems of Gastrulation: Real and Verbal. BioScience 26(1): 36-39
Mlodzik, M. (2002). Planar cell polarization: do the same mechanisms regulate Drosophila tissue polarity and vertebrate gastrulation? Trends in Genetics, 18(11), 564–571. doi:10.1016/s0168-9525(02)02770-1
Raff RA (1999) Larval homologies and radical evolutionary changes in early development. In: Bock GK, Cardew G, eds. Homology: Novartis Foundation Symposium 222. John Wiley & Sons (Chichester) pp 110–120

Ah, yes, Bio-Complexity, the silly pretend “journal” that has 31 editors and managed to publish a grand total of 3 articles in the calendar year 2022.
Whenever a journal has 10 times more editors than annually-published articles, something is definitely wrong.
And then those 31 editors let this piece of crap slide through.
Haeckel himself seemed to have recognized the thorny problem of early embryonic adaptations:
https://en.m.wikipedia.org/wiki/Caenogenesis
Excellent dissection of Creationist Conflationary Confusion. Short version may be summarized: Deuterostomes have a dorsal central nerve cord whereas Protostomes have a ventral dorsal nerve chord. Contrary to introductory textbook orthodoxy, this may be the only real distinguishing feature between the two. It would be interesting to determine which was more ancestral.
Excellent dissection of Creationist Conflationary Confusion. Short version may be summarized: Deuterostomes have a dorsal central nerve cord whereas Protostomes have a ventral dorsal nerve chord. Contrary to introductory textbook orthodoxy, this may be the only real distinguishing feature between the two. It would be interesting to determine which was more ancestral.
This Swift dude seems to be a data point against the idea of nominative determinism. Unless he can run really fast or suchlike maybe?
StevoR @ 4
He thinks he can read through textbooks in one afternoon and break through the arguments of generations of scholars.
I imagine him sittning there, snorting cocaine and listening to “Roadblox”.
No standards at all? Bullshit. They have the most exacting standards: it needs to sound vaguely sciency and it needs to back up creationism.
Maybe he should try finding evidence that supports creationism, instead. Even if evolution turned out to be a completely wrong theory, that wouldn’t support an alternative. There are a lot of goddist theories of creation – maybe he should focus on proving the buddhists are wrong and the christians are right – something theists have notably struck out at for millennia. “Look not at the mote in your brother’s theory when thine own is a shambles,” or something.
How nice that they practice “responsible quote-mining.” It’s so shallow and ecologically safe! Never very deep and completely frack-free.
As usual, it’s hard to tell whether Swift is stupid or blinded by his faith or both.
That’s because no one imposes standards on them, or punishes them for violating them.
Don’t these idiots have something better to do than wasting time dwelling on claims that are easily debunked time and time again, or are they comfortable with the old Nazi tactic of spewing out repetitive lies long enough for gullible and naive people to believe them?
OT
In case you want to rest your brains after this steaming pile of xxxx, the latest God Awful Movies has arrived at Youtube. They dug up a documentary claming all disease is caused by root canals- we have found a cerebral rival to David Swift!
I found a real article, with conflicting evolutionary pressure;
Variation that protected against Black Death helps against respiratory diseases but increase autoimmune disease risk.
https://medicalxpress.com/news/2023-03-variation-black-death-respiratory-diseases.html
OT
Lie detection tests have worked the same way for 3000 years and they are still rubbish.
https://phys.org/news/2023-03-years-theyre-hopelessly-inaccurate.html
Once again I learn something new from Pharyngula, I had no idea early stages varied so much – thank you PZ.
I fail to understand how the creatidiot thinks the variation is evidence that god did it.
I don’t think “scientific creationism” is intended to persuade anyone. It’s just so they can reassure their followers that they have their own “science” to refute the evil secularist scientists. As with climate change denial, the veneer is enough to persuade people who want to be persuaded and don’t really understand the details.
I got a strong sense of déjà vu reading this. Isn’t this exactly the same sort of purblind quote-mining, selective reading, willful misinterpretation, and so on and so forth, of the actual scientific literature, as blarphed out by wossface, Jonathan Wells (and maybe others?), way back in the mid-oughts, that you responded to in the exact same way more than a 15 years ago? I even had in the back of my mind: “Isn’t this where that developmental hourglass thing goes? . . . Oh, aha, there it is!”
History fricken repeating…
@9.imback : Both.
@ 11. Owosso Harpist : Yes.
Is there yet any functional explanation of why Raff’s ontogenetic “hourglass” exists? Intuitively, one would expect that once divergence between developmental pathways happens, it would continue. There must presumably be some selective pressure constraining early developmental paths to lead to the “phylotypic” stage (I take note that there are still significant differences even between closely related species in this stage) while allowing and indeed presumably promoting changes in early development that do not violate this constraint, but what is it about the phylotypic stage needs to be preserved?