
Once upon a time, deep in the Precambrian, this was the planet of worms. Well, actually, this was, is, and always will be the planet of bacteria, but if you filter your perspective to just organisms above a particular size, and if you’re an animal writing about it in the modern day with a chauvinistic attitude that allows you to ignore that it was also a planet of algae, that would become a planet of plants, on a world that also is built of soil formed by lichens and saturated with fungus…if you ignore all that, OK, it was a planet of worms.
Late in the Precambrian, the oceans were full of accumulated muck and it was a good time to be a worm — a slender, plastic body, able to burrow and plunder the detritus of it’s nutrients, digging shelters or writhing in the freshest debris up top. Worms were everywhere, and the family was diverse. There were hairy ones, thick ones, slender ones, spiky ones. There were worms beginning to assemble bits of armor, either extracting calcium from their environment or crosslinking stable sugars to create chitin, because there were also worms that were developing the habit of eating other worms, rather than farming the muck.
Worms were and are diverse. One successful subfamily of worms were segmented — they had begun to evolve more elaborate mechanisms of specializing body parts, so instead of just a simple tube, they had ways of setting aside regions for essential functions, like a head end for eating, a tail end for excreting and reproducing, and a means of making lots of repetitive copies of subunits of their bodies. So, for instance, when they evolved a way to make a bristle at one place, it would be repeated in a whole chain of bristles down the length of the body.
Now as I said, these worms were diverse — they’d been proliferating and diverging for hundreds of millions of years, and each new kind of worm created a more complex environment that permitted even more kinds of worms to flourish. Different lineages of worms had different histories and different sets of attributes. At the end of the Precambrian, not only had they fueled their own diversification, but they’d also changed the planet. They’d stirred up long-buried nutrients and brought about a population explosion, and other organisms (you know, those neglected algae) had been changing the atmosphere, and it was a different world, ripe with opportunity for even more novel forms and opening up new environments, and there were worms fortuitously endowed with traits to take advantage of those opportunities.
In this vast family of worms, I’m going to mention just three cousins — three Precambrian worms that were going to found new dynasties in the Cambrian and on.
One cousin had inherited a couple of extraordinarily useful endowments. She had limbs. Lots of limbs. She was segmented, and initially had figured out how to sprout muscular appendages from each segment, and so she had an extravagant superfluity of limbs, and each could be specialized in novel ways. She was a walking Swiss army knife. She also had acquired bits of chitinous armor, which would also serve her family in good stead — she was the armored and armed cousin, who would found the great family of arthropods.
If she’d had foresight, she would have smiled wolfishly at the prospects for her descendants, if she’d had the kind of mouth that could smile. Instead, she had a face full of legs, each one already specializing into stabbing needles and gnashing blades and pulverizing clubs, which, come to think of it, is even better than a smile for communicating wolfishness.
The second cousin had not inherited any limbs or armor at all. She was smooth and torpedo-shaped. Her endowment was the beginnings of an internal skeleton, a notochord, which made her a fast and efficient swimmer. She was the mother of chordates, and she was the swift cousin — what she gave to her descendants was the ability to run away quickly. In contrast to her armed cousin, she also had a purty mouth, a delicate basket of rods and cilia and membranes, a pharynx that was an efficient filter feeding tool. Someday, her descendants would be able to smile, but only after radically rearranging that complex endowment.
The third cousin…well, I’m going to call her the humble cousin. She had no arms, either, just one big muscular foot that she could use to crawl about on the bottom of the ocean. She had a gland to secrete a shell, but it wasn’t articulated like that of her armored cousin, so it really was a purely defensive structure — she couldn’t shape it into knives and spears. She also had a talent for mucus, which is not usually the kind of thing to make one popular at parties. She’s not so obvious as the founder of a successful family, but she was. She was the mother of molluscs. She didn’t have much of a face — a sheet of muscle with a muscular sphincter — so she wasn’t much for smiling.
Time passes. Arthropods flourish and take over the world — in the modern day we’d have to say we’re living on the Planet of the Insects, except that that external armor turned out to impose some limitations on size so they never got big enough to support a large brain, didn’t invent the internet, and aren’t writing the story of history. But that doesn’t matter much when they’re the first to invade the land, first to take to the skies, and are filling practically every ecosystem on the planet. Those limbs turned out to be a really lucky and extremely handy if you wanted to interact with the environment in complex ways. Eating, walking, flying…limbs make those behaviors relatively easy, and enable all kinds of specialization.
Limbs do have a limitation — they’re a real drag for swimming, where simple, streamlined torpedos rule. The swift cousin does well in the open sea, and builds a huge clan of fish. Evolution tinkers with them, too, though. Even a torpedo can benefit from fins, so those fish that can manage a little fin-fold swim straighter, and those that can flex a fin become more maneuverable. The heavy duty modifications occur in the pharynx. The bigger the animal, the less useful a delicate ciliated basket is for feeding, so it’s dismantled, rearranged, repurposed. It’s used for respiration; parts of it are beefed up and turned into a grasping clamp, the jaws; it becomes a key element of the blood circulation; its membranes are used for ion exchange, and some of the tissue is specialized for managing salt balance; in fact, the whole front end of the chordate — now vertebrate — gets massively renovated, not just to recycle elements of the pharyngeal apparatus, but to build a bigger brain and a sensitive sensory apparatus.
But jaws don’t quite have the versatility of limbs. It took almost a hundred million years after the beginning of the Cambrian for vertebrates to scrape together a kludge to give some of their members a small set of manipulatory/locomotor appendages. This process involved localizing four patches of epidermal thickening1, reusing signals to recruit mesodermal tissue into the zones, and then recycling signals used for patterning the longitudinal axis of the animal to pattern the length of the limb. It’s an amusing glimpse into the way evolution tinkers to see how vertebrate limbs are such perfect examples of bricolage: a piece here, a piece there, jigger them about and reassemble them to make a protruding limb, with the same Hox genes used to specify the organization of the hindbrain lifted wholesale and used in new ways to organize upper arm, forearm, and digits of the hand.
It’s also a testimony to the utility of limbs that they’ve evolved multiple times. There must be considerable incentive for some lineages to reach out and touch the world somehow, as these projecting bulges keep popping out. But they also reveal the other side of convergent evolution: we can get similar function by very different mechanisms, and internal structure reveals historical differences. Here’s a cartoon of cross sections of various limbs to expose those differences.
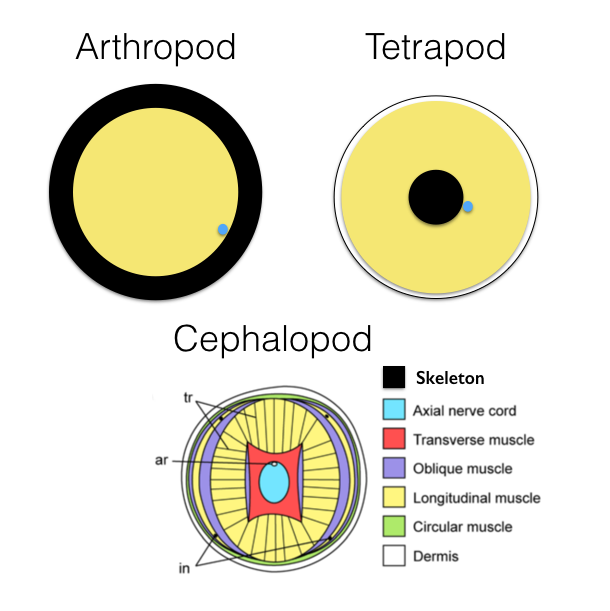
So arthropods had limbs from the earliest times in their history, and are a defining character of the clade. They also acquired an armored exoskeleton made of chitin. When we look at their limbs in cross-section (top left), what we see is a strong, supportive skeletal tube with muscles (in yellow) on the inside. Those muscles are primarily longitudinal — that is, they attach to the inner wall of the tube and extend the length of that tube to attach to the inner wall of the next tube in sequence (or to internal protrusions called apodemes), and the tubes are linked by articulating joints.
The subgroup of the vertebrates, the tetrapods, that finally got around to evolving jointed limbs, have a mechanically different solution (top right). Their limbs have a central bony rod to which longitudinal2 muscles attach, and extend down to the next rod in sequence. Each bone is articulated with the next with joints made of cartilage. Vertebrates and arthropods have come up with roughly similar solutions to movement, constructing rigid structual elements that can be flexed relative to each other by the actions of muscles, but they’re inside out relative to one another. As it turns out, having an internal structural support is arguably better engineering at large sizes, while having an external support that doubles as armor is arguably more efficient at smaller sizes, which is one reason arthropods rule the animal domain below about 10cm in size, while tetrapods are living in lonely majesty above that size.3

But wait! What about that homely cousin, the molluscs? I was wrong to call them homely. They blossomed. The nudibranchs are really the most spectacularly colorful animals in the ocean, and they used a body that, thanks to that muscular foot, is practically a solid wall of intricately controlled muscle fibers in multiple orientations. They have a muscular versatility that puts Terry Crews’ pecs to shame, but one thing they lack is a rigid skeletal framework. No bones or exoskeleton to tug on! All they can do is exert force on other muscles and on the flexible dermal connective tissue, which sounds as if it would be a serious limitation, but then you watch a sea slug dance through the water, and you have to rethink that assumption.
This is where convergence gets interesting: some members of the molluscan clade also evolved limbs de novo. Cephalopods are simply slugs with ambition. They also evolved new limbs, which had to be even more difficult than the path tetrapods took — molluscs lack any kind of articulated skeleton at all, so there aren’t many conveniently rigid structural elements to act upon. So they resorted to a different solution. Tentacles.
The bottom illustration in the diagram above is a cross-section through an octopus arm. There is no skeleton. It’s just layers of muscles, some longitudinal, some transverse, some circumferential, all pulling in complex ways to change the shape of the appendage. The closest thing we humans have to something like that is our tongue, which contains intrinsic muscles that do not have a bony attachment, and which can change the organ’s shape in interesting ways. But look at that diagram! They have all kinds of muscles in all kinds of orientations, and also unlike the arthropod or tetrapod, a robust central nervous organ with chains of ganglia extending the length of the arm. It takes a lot of local circuitry to control something as complex as a cephalopod arm.
That’s three different ways to make a limb right here on our one solitary planet. That’s interesting in both a developmental and evolutionary way. Do all three use similar processes? We know quite a bit about tetrapod and arthropod limb development, and they aren’t homologous at all. Primitively, every segment in an arthropod has a limb; vertebrates had none. As is typical, limbs had to be built using the common molecular toolkit, and we find that homologous genes are used in similar ways. Insects use a gene called decapentaplegic as a morphogen to organize their limb fields. Vertebrates use the decapentaplegic homolog Bone Morphogenetic Protein to regulate skeletal formation in the embryo. We know that molluscs also have decapentaplegic — they use it as a morphogen to control shell shape. Does it get recycled into cephalopod arm formation?
We also know how squishy vertebrate limbs grow. There is an epidermal thickening called the apical ectodermal ridge that secretes a morphogen, FGF to induce proliferation and extension, and another part of the limb bud called the zone of polarizing activity that uses another morphogen, Shh, to induce polarity (the dorsal side of your arm is different than the ventral). How do cephalopod arms develop? Is it similar, with the same kinds of patterns of proliferation and extension, and do they also recruit the same or similar molecules to do the job?
People are always asking me why I’m known for my fascination with cephalopods, when I work on a limbless vertebrate, the zebrafish, and this is why. Because asking why things are the way they are requires asking about the way they are not. Understanding how our biology came to take one particular path should involve looking at all the different paths, and thinking about alternatives. So if you want to really understand vertebrate limb development, it’s important to compare it with invertebrate limb development.
My introduction has gone on way too long, so I’ll post the answers (and mostly, lack of answers) to my questions later. For now, just remember that the issue is how different organisms build a protruding appendage, a very general question, and we want to know more about how a group very different from our own, that has diverged from the vertebrate lineage for at least 600 million years, has constructed their unique version of an arm.
Nödl et al. (2015) The making of an octopus arm. EvoDevo 6:19
1One of the things that always annoys me in SF movies is the ubiquity of bipeds tottering about. That we only have four limbs is entirely a historical accident of our clade — and bipeds like us are the result of a clumsy evolutionary effort to free up a pair from locomotory duties to be more manipulatory. We’re weird! Why do we keep unthinkingly imposing our historical limitations on every alien species we imagine, especially when our own planet is so rich in species that have no such arbitrary constraint?
And don’t use that “movie budget” excuse: we have CGI aliens all over the place, and even there they simply map them onto distorted bipedal forms. We even have movies like Avatar which imagines an entire diverse alien ecosystem with diverse body plans, and then chickens out and turns the intelligent aliens into slender big-eyed anime models4.
2Of course we have some greater complexity than just longitudinal muscles — obliques, for instance, that rotate the radius and ulna, or the layered sheets of muscle that make up the abdominal wall.
3On the topic of imaginary aliens — use our extant patterns as a guide. Imagine an equivalent Precambrian world elsewhere, where one group gets a mix of traits, a complex oral structure plus an exoskeletal armor, whie another group gets a different mix, multiple segmented limbs plus an internal skeleton. Work forward. Later, you’ve got small heavily armored snake-like things with intricate mouthparts, and giant fleshy beasts with limbs sprouting all over the place.
And of course, the creatures with all the feet evolve into galactic imperialists, conquering the universe to accumulate enough real estate to house all their shoe stores.
4Also chickening out: I liked the diversity of limb structure in the John Carter movie, but once again, the hero species defaulted to fully humanoid, to the point where a human would find them sexually attractive. Which is bizarre — the Red Martians in Burroughs’ stories lay eggs. Somebody needs to write the story of John Carter and Dejah Thoris’s wedding night, when the wife disrobes and reveals to her husband…a cloaca. Does the sexual attraction endure?

Is this for your next book?
I’ll buy it.
No, I won’t be posting anything from the next book here.
If Internet porn is anything to go by, yes.
“One of the things that always annoys me in SF movies is the ubiquity of bipeds tottering about. ”
It’s difficult to get actors who can do 8 legs.
Hire four of them.
A 32-legged SF critter? That could work.
Worse, every vertebrate(ish) Pandoran species depicted in AVATAR other than the Na’vi are hexapodal and have two interface tentacles. The Na’vi have only four limbs and one interface tentacle. I find it highly unlikely that the Na’vi evolved on that moon.
I had once imagined an extraterrestrial creature for a sci-fi story I never wrote. They were multi-limbed invertebrates, but they had learned to manipulate found materials; sticks, bones, branches, etc, to use as stilts so they could move more efficiently out of water. Eventually, they built inorganic jointed skeletons they could wrap their boneless bodies on, and by that means, built a star-faring civilization. When encountering humans, they were able to re-engineer their skeletons into a tetrapod format, and by way of their octopoid chromatophores, could pull off a passable imitation of a human, as long as nobody looked too close….
That was an excellent story, thank you.
When it comes to aliens, anyone looking to make a movie or TV show with aliens should read the Sector General books by James White. He grokked alien.
Although the newest War of the Worlds was a crappy movie ( it should have been called Tom Cruise vs. The Aliens) the aliens in it had an interesting body plan. They were pentapods that walked on 3 legs. A rear leg and 2 forward leg/arms that were also used as hands. They also had 2 small forward arms that would work with the leg/arms to hold things and also, presumably, for things requiring fine motor skills. A fair amount of thought and skill went into designing them – its too bad the writers completely missed the point of the original story.
In Mission of Gravity, Hal Clement had an interesting, arthropod derived (possibly) alien race adapted to a high gravity planet. Their emotions seemed rather human-like if I remember correctly.
I’ve always thought that Robert L. Forward had a really unique alien species in The Dragon’s Egg: sentient beings living on the surface of a neutron star.
http://www.amazon.com/Dragons-Egg-Del-Rey-Impact-ebook/dp/B004G8PJDA/ref=sr_1_1?s=books&ie=UTF8&qid=1452111726&sr=1-1&keywords=dragon%27s+egg
This is great, PZ!
More. please!
My head-canon for the Na’vi is that they were created en masse, complete with fictional history, by the real rulers of Pandora as a response to the human invasion, probably using captured H. sapiens as templates.
What about Jabba the Hut — a large, slug-like creature with arms, but no legs.
This is why I keep coming back to this blog. The article is fascinating, and then the commenters make me smile (especially #’s 3 & 15).
Great post PZ.
And yeah, given the vast variety of life just in this planet, is unbelievable how narrow aliens are represented.
Besides a lack of appreciation for Earthling diversity, I wonder if it’s also because human-like aliens are more relatable.
I wonder, rather than the differences, what could be the *necessary* similarities?
I have hard time imagining a non-social species to build a civilization, so maybe it’s a requirement.
If you are social, you also need a way to coordinate and share information with other individuals.
You also need a way to manipulate the environment, through specialized limbs or especially tight collaboration.
Probably the individual needs to be able to learn for the longer term.
I wonder if there are ways to do without any of these.
Charming.
But, of course. Otherwise we wouldn’t have attempts of seduction of Capt. Kirk by females of various hues. And 50+ years ago I decided that I would suspend my disbelief about the reproductory mechanics of John Carter and Dejah Thoris just as I ignored the incompatibility of Sarek and Amanda. Complex relationships between the genders is such an easy plot device.
Great post PZ!
Ahhh. The John Carter of Mars books. My dad was a huge Burroughs fan. And managed to pass it on to me and my sister.
One of my dads favorite stories was one of his brothers submitting a book report in school on one of the books. Been a long time since I read them and I don’t remember which book Dejah Thoris layed her’s and Carter’s egg. That’s the one he wrote the book report on. Got sent to the office and had his mother called in. This would have been in the mid 1920’s…
Oh. And that was really fascinating PZ. Thank You!
@Tristan #7:
They were intelligently designed?
You mentioned the tongue as a vertebrate’s version of a tentacle. Another example is an elephant’s trunk.
Lovely as tentacles may be, I think they have certain disadvantages compared to limbs with skeletons, whether endo- or exo-. (I am no expert on biomechanics, and I welcome correction from anyone who is.)
First, tentacles aren’t very good at resisting compressive forces along their length. I know of no animals of appreciable size that can support their weight on tentacles. An octopus may walk on his tentacles, but that’s underwater, where buoyancy provides most of the support. Plop him on the deck of a boat and the best he can do is slither toward the gunwales.
Second, I suspect that slender tentacles are inherently much weaker than fingers, and therefore wouldn’t be as useful for a toolmaking species. This notion requires some explanation:
As I understand it, tentacles work largely as muscular hydrostats. Cell membranes divide the muscle into many smaller compartments, each of which has (on a short time scale) a constant volume. When a muscle cell contracts, it becomes shorter in one direction but fatter in the others. By clever coordination of all the shortening and fattening, a tentacle can change its shape even without any rigid members to attach to.
However, this arrangement means that the when a tentacle bends, the force is generated pretty much at the site of the bend. For the tentacle to be strong, it has to be thick, to make room for enough muscles.
Fingers, by contrast, are powered by muscles remote from the fingers themselves, and connected to them by tendons. The fingers can be slender but strong, because there’s plenty of room on the forearm for bulging muscles.
Conclusion: a species with fingers is likely to be handier with a hammer than one with tentacles — and better at swinging from branch to branch.
That is correct, tentacles won’t be good at handling compression, where a rigid skeleton is much more helpful. They lack good mechanisms for increasing efficiency by storing energy and releasing it in recoil.
As is clear, though, they do have advantages: not strength, but great dexterity and flexibility. They also perform well in water, where buoyancy can compensate for deficiencies in strength. Like anything, they’re going to have both advantages and disadvantages when compared with other kinds of limbs.
So, when one is writing an alternate universe scenario …
is endo really better than exo?
or are there other, weirder, possibilities?
why would waterworld be a barrier to technology?
really
do we really need fire?
thanks, PZ! That first paragraph set the tone for a great read. Bookmarked for my friends who have not been exposed to your prose.
@chigau #28: exo seems to be a liability if you want to grow beyond a certain size, which also /seems/ to be necessary if you want much of a brain.
Basically exo, at least the way arthropods implemented it, runs out of surface area where to hook squishy stuff and seems tightly coupled with their breathing/circulation system.
I would love to learn that some arthropod out there has any outstanding innovation on this.
AFAIK the most “intelligent” arthropods are mantis shrimps (long term memory, long term pair bonding, recognize individuals they have met before).
It’s also noteworthy that having an endoskeleton with articulated limbs has served cetaceans very well. Heck they managed to feel a few niches without even being able to breathe underwater.
I wondered myself about underwater civilizations.
My interim explanation is that it’s difficult to source materials for tool making that can withstand the environment for long time.
Hey, maybe there WAS an underwater civilization but they died out leaving no fossils and no tools that could last long enough.
@PZ Myers
Futurama did a joke like that. The main character (Fry) falls for a mermaid, considers marrying her . . . and then it turns out they don’t have sex the way regular humans do – she’s confused at his genitals when they get into bed together and tells him he’s supposed to spray something over her eggs.
Forget about her cloaca. What will her reaction be to his penis? And I’m not even going to think about what happens when she confuses his anus for a cloaca. Then again it would make a great script for a porn movie and there is probably someone somewhere warped enough to make it.
Damn but your students are lucky, PZ. They get to be exposed to your exceptional teaching skills all the time!
Thank you for giving this to the non-specialist/non-scientist audience too, it’s great science writing – intriguing, easy to grasp overall without anything dumbed-down :-)
(wish my mum were still alive to send it to – a developmental biologist (mostly research, some teaching), she would have loved this)
Thanks for mentioning this, Caine – I picked up Hospital Station in a charity shop many, many years ago and really enjoyed it, but I didn’t realise until today that it was just the first of a series. I’ll have to see about tracking down the others now.
Do not taunt Rule 34!
Dunc @ 34:
The whole series is now available in e-book form, if you have a reader. I’ve never come across any other books that do aliens as well as White. While he was extraordinary when it came to aliens, he wasn’t so great when it came to women – the sexism is of a particularly stupid variety, and will cause near-fatal eyerolls. Other than that, the stories are really excellent (he doesn’t focus on women much at all, so thankfully, you don’t have to put up with continual sexism.)
“the creatures with all the feet evolve into galactic imperialists, conquering the universe to accumulate enough real estate to house all their shoe stores”
Ah, the Shoe Event Horizon! You get a summary of Douglas Adams’ theory of evolution and disaster economics in The Restaurant at the End of the Universe, but it’s far better on the radio – the script is at that link. The economics lesson:
https://www.youtube.com/watch?v=nEI19kJ5GfU
and Hig Hurtenflurst, the risingest young executive in the Dolmansaxlill Show Corporation, is definitely a galactic imperialist, with a Shoe Shop Intensifier Ray, Foot Warriors whose feet are the wrong size for their shoes, and his spelling of ‘revoked’ as ‘k-i-l-l-e-d’.
It has only been recently that CGI would allow a real variety in possible alien configuration, but that would remove the whole romance thingy. Then there is this:
@10: As it happens, I literally just finished reading Hospital Station again. It’s a good read, but Conway’s superhuman ability to grok exactly what’s happening before anyone else kinda puts me off. James White’s aliens, and his system for classifying them, is AWESOME, though!
Re : #37
In Babylon 5, Centauri males had six…
With a hexapodal evolutionary history, they could have given the Navi WINGS! Massive missed opportunity for awesome, imho.
One of the arthropods’ trade offs wrt their exoskeleton and size is the need to molt to grow. Not only is the organism armourless and vulnerable during that transition, but they also lose their support against gravity outside of water for the period.
So how do echinoderms fit into this?
(I have to admit, from the title, I was hoping this would mention the trichords. I played SimEarth as a kid.)
Yes, as far as number of individuals is concerned, but how about by weight? I would have guessed it’s the planet of plants on that criterion.
Great article! Also, I love the discussion of sf aliens in the comments.
Personally, I’m a sucker for the truly alien aliens that populate the work of Stanisław Lem. The proto-sentient ocean of Solaris, the symbionts of Eden, the… stuff in Fiasco, the signal-senders in His Master’s Voice, and so on. (In a sense, he was merely using the trope of sf aliens in order to play with the meaning of sentience, intelligence and communication. This is clear because of his other work where the same questions are explored by other means, for instance using AI as in The Cyberiads, Mortal Engines, Golem XIV.)
The only other aliens I’ve found that compare to Lem’s for sheer alienness are those in Peter Watts’ excellent Blindsight and its sequel Echopraxia. Even the future humans there have drifted quite far from something we can comfortably identify with.
For the biological and sociological invention, I adored the “puppies” in Vinge’s A Fire Upon the Deep (telepathy and collective minds with a plausible biological mechanism!) and his “spiders” in A Deepness in the Sky (arthropod industrial revolution!), as well as Samuel R. Delany’s three-sexed evelm and mysterious Xlv in Stars in My Pockets like Grains of Sand (sex with aliens!).
From the older classics, I fondly remember the above-mentioned flat arthropods in Hal Clement’s A Matter of Gravity (physically plausible intelligent life in super-high gravity!), the beautiful depictions of the Jovian fauna in Clarke’s Space Odissey saga, and also the energy beings form another universe in Asimov’s The Gods Themselves.
For those who liked A Matter of Gravity, Greg Egan’s Incandescence is a quite similar read and is even better at the hard physics. I haven’t read it yet, by I hear that Greg Egan also did a wonderful job with his Orthogonal trilogy, where he built his really truly alien beings starting from the alternative laws of physics up — the ultimate hard sf aliens?
On the other hand, although I love his work for many other reasons, I always found all of Iain M. Bank’s aliens to be too naïvely anthropomorphic, at least mentally. Even the Minds — awesome as they are — have suspiciously human emotions and values.