What a cute little dead baby.
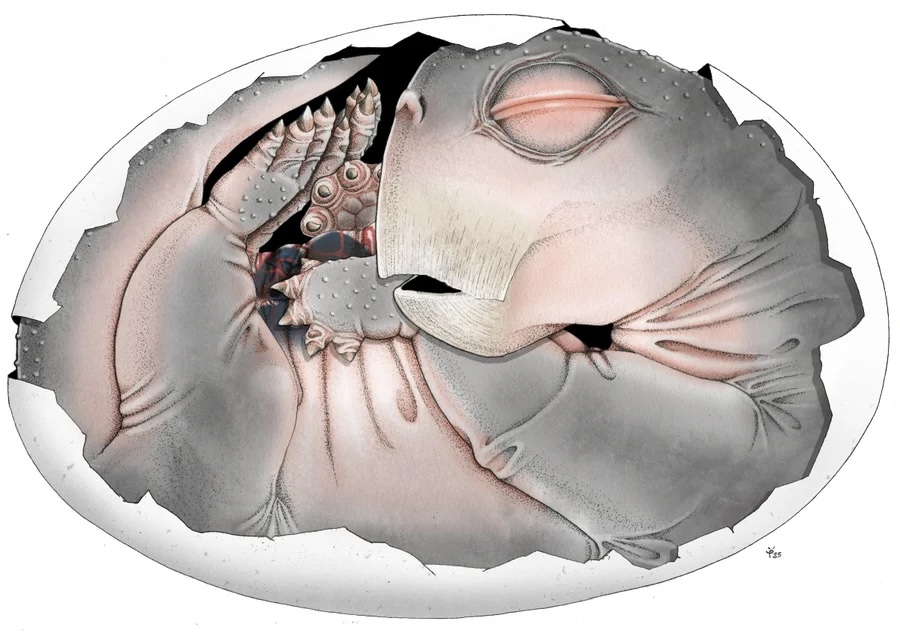
That’s a Lystrosaurus embryo. If you don’t know Lystrosaurus, it’s an amazing species that survived the Permian extinction and experienced a remarkable population boom — it was a large vertebrate that came to dominate the planet after that mass extinction, yet most people have never heard of them. We ought to pay more attention! Memento mori, and all that.
Anyway, this fossil answers the least surprising question ever.
Detailed imaging of a 250-million-year-old fossil has revealed the first proof that the ancestors of mammals laid eggs. The discovery answers a long-standing question about the reproductive biology of our ancient forerunners and hints at how they managed to flourish in the aftermath of the biggest mass extinction in Earth’s history.
Scientists have long assumed that the ancestors of mammals—a group known as the therapsids—laid eggs like today’s platypuses and echidnas do. But they lacked any direct evidence of synapsid eggs in the fossil record.
It’s good to be able to tick off that one box, confirming that Permian therapsids laid eggs, but it’s hardly news. I thought this was much more interesting:
Most importantly, the new images reveal that the two halves of the lower jaw had yet to fuse in the youngest Lystrosaurus specimen. In turtles and birds, the lower jaw fuses before birth, allowing the baby to feed itself after hatching. The unfused lower jaw of this Lystrosaurus is therefore another indication that the animal died while still in its egg. The other two specimens exhibit signs of having been somewhat more mature; the largest one was preserved in a splayed-out posture that shows it was not in an egg and had traveled some distance before dying.
Mammalian ancestors invested more in maternal care than some other organisms. That might be a clue to how our clade survived through a couple of mass extinctions.