I’ve only just noticed that I have a fondness for food metaphors when talking about development — gastrulation is a peculiar way to make a jelly sandwich, neurulation is like rolling up a burrito, and somite formation is a meatball sub. They sort of illustrate the arrangement of the tissues involved, but of course they all have shortcomings…but then explaining how the metaphor doesn’t work can be just as informative as the metaphor itself.
For instance, early in its development, the vertebrate embryo consists of two epithelial sheets, the epiblast and hypoblast, pressed against each other like two slices of bread. That’s easy to visualize. It also allows me to explain the core idea of an epithelium — a layer of cells tightly linked to one another to form a continuous more or less two dimensional sheet. A lot of animal development is about epithelia folding and contacting other layers. But another important concept is that some cells are not in sheets — they’ve dissociated and are moving in a loose mass surrounded by an extracellular matrix. This is called mesenchyme. Mesenchyme would be the gooey jelly between the two sheet-like bread slices.
But here’s where the metaphor falls down: this jelly sandwich is made by a portion of the bread slices folding inward — invaginating — or falling apart — delaminating — and migrating into the space between the bread slices. This is clearly not how we make jelly sandwiches. But even that weird difference is important and illuminating. In addition to coherent sheets forming tissues and organs, another hallmark of animal development is the disassembly of sheets into masses of migrating cells. This process is called the EMT, or epithelial-mesenchyme transition, and it’s not just important during development, but is also invoked again by cancers when they metastasize.
Here’s another deviation from standard human kitchen practices: after the jelly has formed, part of the bread is rolling up into a tube to form the nervous system (the burrito!), and simultaneously, that jelly is organizing itself into a series of balls — it’s also making a meatball sub. If you are finding this concurrent mix of odd sandwiches unappetizing, that’s OK — don’t eat any embryos, then.

But this is another of the extremely cool processes that is going on during the pharyngula stage of development. First we have cells undergoing EMT and leaving their epithelium of origin, migrating into the interior of the embryo, and then a significant subset of these mesenchymal cells clump together to form these ball-like masses called somites that are organized in an orderly row down the length of the animal. Somites are relatively undifferentiated balls of mesoderm. Let me emphasize that: they are not muscle, or bone, or connective tissue, common mesodermal derivatives, at least not yet. They are blobs of condensing mesenchyme.
It’s the next series of steps where they begin to depart from my metaphor even further. The somite begins to differentiate regionally — parts of it are going to follow new paths of development. First, a portion of the myotome ventrally and medially begins to fall apart and stream towards the central notochord. This part is called the sclerotome, and the part it leaves behind is the dermomyotome. The sclerotome is going to differentiate into bone, particularly the vertebrae that surround the notochord and spinal cord. So eat your meatball sub fast, before a quadrant of the meatballs turns to bone!
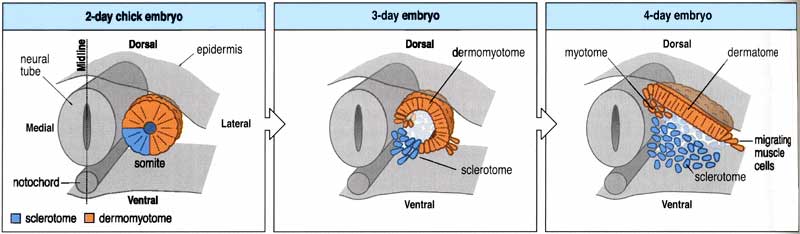
The formation of the sclerotome, by the way, is an example of induction: signals from the notochord tell nearby cells of the myotome that their fate is to become bone. We can do experiments in which additional pieces of notochord are transplanted near the myotome, and they will recruit more cells to become sclerotome.
The dermomyotome is going to divide as well to form a tissue called the dermatome, which will contribute to the connective tissue of the skin, for instance, and another part called the myotome which will form the muscles proper. The myotome is just the start of a long series of developmental decisions that will transform an undifferentiated mass into the complex collection of varied cell types that will make up the trunk and tail of an animal, and that will get drawn out and shaped to form the limbs of tetrapods.
You might be thinking somites are amazingly cool, but if you’re like me, you’re also wondering where they came from — can we say much about the evolution of this pattern of organization? And the answer is, only a little. This way of building a body is deeply primitive to all chordates, so all extant species are doing something very similar.
The tunicates are an exception. Tunicates have definitely gone their own way — they’ve thrown away segmentation, for instance, and just make a continuous mass of contractile cells along the length of the tadpole larva. They’ve also dismantled the regulatory linkages between their Hox genes, and those genes are no longer in a tidy colinear sequence in the genome. So they’re interesting in a unique way, but they are highly derived and do not reflect the ancestral condition.
Amphioxus, on the other hand, is phylogenetically more distant from us than are the tunicates, but does form somites — which tells us that the ancient pre-Cambrian worm, our common ancestor, almost certainly built themselves from the meatball sub recipe. But Amphioxus and vertebrates also exhibit some interesting differences.
For one, Amphioxus doesn’t make myotomes by balling up a loose mesenchyme; instead, it’s all epithelial. They form from an evagination of other epithelia, and retain their epithelial character — so they aren’t jelly-like at all, but are sheets formed by folding in another sheet. Some cells do delaminate and contribute to connective tissue, though, so they are capable of EMT, they just do it to a much more limited degree than seen in vertebrates.
Another question: Amphioxus does not have a bony skeleton; that bone thing is an innovation by early vertebrates. Much of the vertebrate skeleton (but not all!) is formed by sclerotomes; do Amphioxus myotomes have a homolog to the vertebrate sclerotome? Unsurprisingly, they do, and rather than being a mesenchyme, it is once again an epithelium that extends and wraps around the myotome.

Summary of the development of amphioxus non-myotome somite derivatives in embryonic (A), larval (B-E) and adult (F) stages. Note that arrows indicate a change in the relative position of tissues, but it is not known whether this is achieved by active migration and/or differential growth of somitic or surrounding tissues. Abbreviations: ECL, external cell layer; EP, ectodermal epithelium; FBM, fin box mesothelium; MS, myoseptum; MSC, myoseptal fibroblast cell MY, myotome: NT, neural tube; NO, notochord. Stage abbreviations: neur, neurula; GS, gill slit; post-met, post-metamorphic juvenile.
The relationships of all the tissues are the same in vertebrates and protochordates, which tells us the last common ancestor of all chordates almost certainly had segmentally organized myotomes, and a layer of cells set aside to form sclerotomes, even in protochordates lacking a bony skeleton, and that this layer would later be coopted for building bony vertebrae. It also tells us that that last common ancestor had a mechanism for the EMT, even if it wasn’t as widely used as it is in us vertebrates. And it also means we can readily compare the development of the trunk and tail in all chordates.
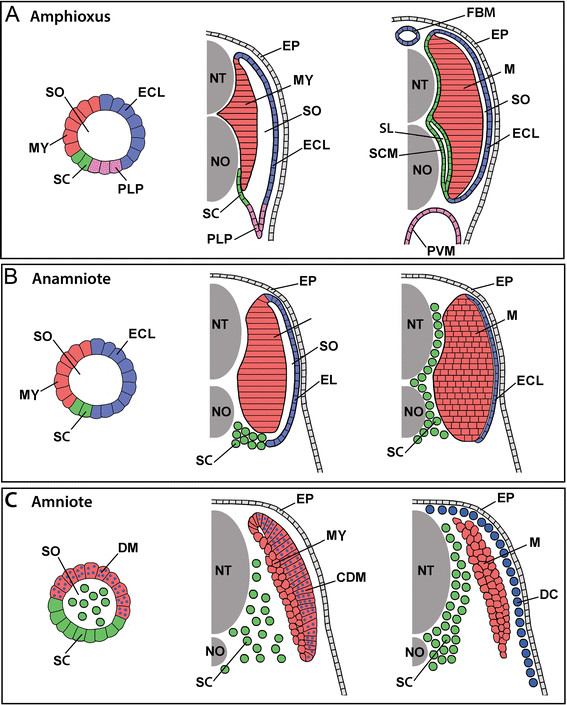
Comparison of somite development and somite compartment derivatives in (A) amphioxus, (B) an anamniote vertebrate, and (C) an amniote vertebrate. For each, somite organization is schematized at early (left panels), mid (middle panels), and late (right panels) developmental stages. The schematics are shown unbent from their true chevron- or W-shape and for simplicity omit the ribs and ventral muscles (and in anamniotes, the myoseptal cells), which are derived from the somites and migrate ventrally into the lateral plate mesoderm. Other details are also omitted, including the difference between epaxial and hypaxial musculature and between phasic and tonic muscle fibers. Abbreviations: CDM, central dermomyotome; DC, dermal cells; DM, dermomyotome; ECL, external cell layer; EP, epidermis; FBM, fin box mesothelium; MY, myotome; NT, neural tube; NO, notochord; PLP, presumptive lateral plate; PVM, perivisceral mesothelium; SL, sclerocoel; SCM, scleromesothelium; SO, somitocoel; SC, sclerotome; M, trunk muscles.
So there was a primeval meatball sub.
Another point, on the side. You may recall a while back that there was a lot of bluster from Islamic creationists about the Quran’s supposed inerrancy of its description of embryonic development. I had this text quoted at me multiple times.
We created man from an essence of clay, then We placed him as a drop of fluid in a safe place. Then We made that drop of fluid into a clinging form, and then We made that form into a lump of flesh, and We made that lump into bones, and We clothed those bones with flesh, and later We made him into other forms. Glory be to God the best of creators.
I argued with Hamza Tzortzis that his developmental chronology made no sense. Note that Mohammed is claiming that first bones are made, and then they are clothed with flesh, but that’s not right. Myotomes assemble, and then split into sclerotome (bone) and dermomyotome (flesh), two tissues that then develop concurrently. Assigning a sequential order to them is a classic “not even wrong” assertion. I think now you can understand why I was confused about how to even begin addressing his argument.
There’s an impressive amount of very thorough work in this freely available paper:
Mansfield JH, Haller E, Holland ND, Brent AE (2015) Development of somites and their derivatives in amphioxus, and implications for the evolution of vertebrate somites. EvoDevo 2015, 6:21.

And here I thought from the title I was going to get a joke about baby eating atheists.
I’m flattered.
I find all this to be very gross. And then, I hear there are actually people called “doctors”, who sometimes deal with all this stuff, and even make money from it?
Unacceptable. This whole “development” process, if that really is its name, simply must be stopped. And what’s more, on certain days, most people agree with that.
Who do we see to get this done?
(Just kidding)
Hmm, to me that seems to be just your average style of wrong.
“Do fetuses have a bony-but-fleshless stage of development?”
“No.”
Aw, you mean that embryos don’t spend their entire time looking just like smaller versions of newborn babies? Surely the anti-abortion crowd wouldn’t lie to further their political goals?
How an embryo is like a meatball sub: they both go down easy with an ice cold beer to wash them down.
There’s good eating on a fetus!
How an embryo is like the cow that was butchered to make the meatball sub
Down here in good ol’ Florida, I keep seeing billboards and bumper stickers, breathlessly informing us that
And every time I see this “argument,” I think: Yeah, it does. So does slaughtering a cow. So what? Neither one has much of a brain or a personality. In neither case are you killing a person. Only the religious could see more worth in a first-trimester human embryo than in a full-grown cow.
Thanks for the post. This is the whole reason I began reading Pharyngula several years ago.
Now I’ve got to run out and get a meatball sub. Do babies taste that good, or would that be too late in the development process? (It probably surprises you that an atheist wouldn’t know the answer to that question, since eating babies is part of the initiation into atheism. Don’t let anyone else know, but I sneaked in the back door.)
A human embryo at this stage would be tiny, gooey, watery, and would probably have a faint (they’re tiny!) flavor of a blood-streaked oil drop. Not recommended.
I was hungry before I read this, but your article made it worse. I could go for a meatball sub right about now or any non-metal sub, really. One step closer to eating a baby, like proper atheists should, eh?
Time for lunch!!
Great post. I’m particularly intrigued by the last sentence of the article: “It will be interesting to uncover the genetic changes necessary to derive a cartilage-producing cell population within a connective tissue somitic compartment. ”
How much do we know about the genes that control all of this? Do we even have an estimate of the number of genes involved?
I really enjoy this subject, especially the multiple ways various animals have evolved a myriad of body plans. I am a little confused by the way the cephalochordates are being assumed to be the ancestral body plan. I thought that within the Deuterostomes the genetic and cladistic analysis resulted in four major groups. Echinoderms, Hemichordates, Urochordates, and Chordates, with Cephalochordates being an oddball within chordates.
Perhaps I am misunderstanding this sentence, but I see no reason to assume the most basal somite was a soft bodied animal considering the great age of the echinoderm family . An interior hydrostatic test could just as easily be a feature of the most basal somite. Is it known which gene sequences Echinoderms use to make stereom, and are they analogs or homologous to the way vertebrates make bone?