The latest fatuous obsession by Paul Nelson, Philosopher of Biology at the Discovery Institute, is a real corker. He has decided that nematodes could not possibly have evolved, because scientists (real ones, not creationist pseudoscientists) have produced an extremely detailed literature documenting their development; because Brenner, Horvitz, and Sulston (no creationists among them) won the Nobel Prize for their work describing the cell lineages to produce the worm; and because he doesn’t understand developmental biology at all. I’ve got palm impressions in my forehead from smacking myself so many times while watching this terrible little video.
This is why Paul Nelson is laughed at by developmental biologists. He cannot be taken seriously. I figured, though, that if you’re not already familiar with concepts and details of development, you might find him credible — he’s so pompously earnest! — so I thought I’d explain all the ways he goes wrong.
First, though, a nitpick of an annoying detail before I go on to the more substantial problems. Back in the day, I did my graduate and post-doctoral work using lineage analysis tools to study the development of the nervous system in zebrafish, and so I’m rather familiar with the history of the techniques. This statement is just wrong:
For the first time, in the history of biology, we were able to see and track the development of an animal, from one cell to the adult. That had never been done before.
Nope. People have been tracking cell lineages in embryos for over a hundred years. We’ve been using chalk particles, carbon, inks, dyes, and natural color markers in embryos to track cell fates for at least that long — does no one remember Edwin G. Conklin and his gorgeous studies of ascidian development?
The nematode work is notable for the detail, and for the fact that nematodes had a remarkably invariant pattern of cell divisions that meant the observations in one animal were precisely reproducible in another. The nematode Nobel was awarded for “discoveries concerning genetic regulation of organ development and programmed cell death” — what made it stand out was that the investigators did genetic and cellular experiments to uncover the mechanisms underlying the pattern.
OK? A minor point, but indicative of how sloppy and shallow Nelson’s understanding is.
Here are the deeper problems that jumped out at me.
The I can’t imagine how this evolved
argument. This is every intelligent design creationism argument ever. First step: find something biological that is really complicated (this is easy, it’s all complicated). Second step: express how impressed you are with the beautiful science here. Third step: appeal to the ignorance of all the non-scientists by declaring that this intricate structure could not have possible arisen by pure chance. Never mind that every part of this amazing biological wonder you’re praising was worked out by scientists who understood evolution.
That’s all Nelson is doing here. We have this complex spatial arrangement of parts in the adult organism, and to that he adds the fact that it is assembled by a complex sequence of developmental interactions over time, and presto! It’s awesomely complex, therefore it must have been designed.
This whole strategy is a cheat and a science stopper. Every level of understanding we add to the process makes it automatically more complex, therefore the more we know the more the ignorant people get to proclaim it’s too difficult to evolve, because as we all know, in the creationist community the rate limiting factor in biological functions is the comprehension of the dumbest bible-thumper in the group…anything above that is an act of God.
But, you know, people in the 19th century knew nematodes as microscopic worms, and would have argued that the existence of tiny squirmy wormies was evidence of god. Then when the tissues of the worm were analyzed by microscope, when every synapse of the nervous system was mapped with the electron microscope, that was declared to be proof of God’s hand. And now we can tease apart the molecules involved in signaling between cells at the 4 cell stage, which adds a whole new layer of complexity to the story, so hallelujah! It must be the work of god!
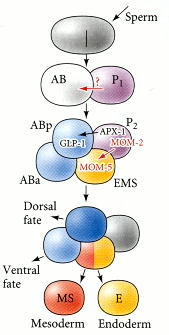
Cell-cell signaling in the 4-cell embryo of C. elegans. The P2 cell produces two signals: (1) the juxtacrine protein APX-1 (Delta), which is bound by GLP-1 (Notch) on the ABp cell, and (2) the paracrine protein MOM-2 (Wnt), which is bound by the MOM-5 (Frizzled) protein on the EMS cell.
It’s a cute trick, to turn every scientific confirmation of the natural, mechanistic processes underlying the development of a worm into a testimonial to supernatural forces. But it doesn’t hold up.
(I must also point out that every person with a passing familiarity with evolution and development will read that figure caption and recognize old friends. Notch, Wnt, and Frizzled are ubiquitous in flies, frogs, fish, and people — the worm is using tried and true molecules that evolved at least 600 million years ago.)
The false syllogism: It’s complex, and specific, therefore it must be designed.
I can tell why Nelson loves the worm example. It’s complicated, which is number one on the creationist list of popular phenomena, but it’s also extremely specific. The worm goes through an exceptionally stereotyped pattern of divisions and cell fate decisions to produce the adult form. This system meets their mantra of specified complexity
, an argument that rests on the elision of the meaning of two words, specific and specified.
The divisions of the worm are precise and reproducible. Every C. elegans makes that first division into two identifiable cells, one called AB and the other P, and we can also predict how AB and P will subsequently divide, and where their progeny will end up in the adult. It is specific in the sense of being precise, accurate, consistent.
“Specified”, as they use it in the term “specified complexity”, means something entirely different. It means built to a plan or specification; it implies a pre-existing blueprint that lays out the instructions to produce a desired end result. In the case of intelligent design creationism, it attempts to turn an assumption into evidence.
In simpler words, they look at a worm and declare, It looks like it was built to a plan! And a plan can only be created by a planner, therefore it was designed.
OK, then, show me the cosmic blueprint that predefines how a worm is supposed to be built, and then maybe I’ll consider the possibility that it was built to a specification.
The lack of an alternative prediction. I’m used to the idea that if you present a hypothesis to explain a phenomenon, you should be able to clearly state or predict what outcome you would see if your hypothesis is false, and what you should see if it is true. Here Nelson points to the stereotyped lineage of C. elegans, where every cell in the adult has a well-defined pedigree, and essentially says A-ha! This could only happen if it were designed!
OK, then, what would it have looked like if it weren’t designed, but had evolved?
If precision pedigrees are the signature of a designer, what about all those animals with less rigid lineages?
I mentioned that I studied neurons using lineage tracing methods. We would have killed for a predictable lineage in zebrafish. We’d go in and inject dyes into cells that we hoped would end up labeled in the neurons of the spinal cord; we had a rough knowledge of what layer of cells were likely to end up there, so we’d poke a random cell in about the right place, wait a day, and then look at where all the progeny of that cell ended up in the embryo.
We had a 2-3% success rate.
So fish aren’t as precise as nematodes. They’d throw a patch of cells in the direction of a tissue, and then the cells would later sort out among themselves which one would do what, with a fair amount of variability. They’re nowhere near as specific as a worm.
Don’t even get me started on the early blastomeres: label one cell at the eight cell stage, and there was so much semi-random mixing of cells you’d get something like this the next day:
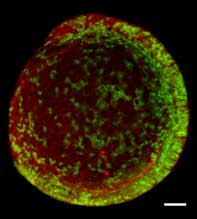
Does that mean fish (and people, who are just as sloppy) evolved, while worms were designed? What exactly are you using as a criterion to differentiate evolved from designed?
That’s actually a rhetorical question, because it doesn’t matter what the organism looks like or how it got that way, they’ll say it looks
designed, which tells you precisely how useless their criteria are.
Development is hierarchical and incremental. In a related point, I have to mention this: the pattern Nelson describes is unsurprising, and fits with everything we know about development in other organisms.
In what is almost certainly a consequence of the mechanisms of gene regulation, development is strongly hierarchical, as we can see in the example of the worm. Cell divisions are accompanied by binary decisions with effects over multiple cell generations. So the worm zygote divides into two cells, AB and P; each of these two cells has different patterns of gene activity that also define gene activity in their daughter cells. The whole is steadily partitioned into subsets, each with a pattern of gene activity that both enables and limits what succeeding generations can do.
Further, this partitioning occurs progressively, in steps, which makes sense since many of the cell fate decisions are determined by local interactions between cells. AB, for instance, looks to the P cell for signals that shape the development of daughter cells. Some of the decisions of nematode cells are autonomous — that is, they will divide in a particular way no matter who else is in the neighborhood — but others are contingent on interactions between cells.
I look at the pattern of cell divisions that have Paul Nelson goggle-eyed and wondering how do it do that, and I’m seeing an entirely predictable general pattern, derived from known mechanisms.
But I’ll tell you an observation that would have me baffled and wondering if maybe there was a Designer after all: if the nematode cells divided with no progressive differentiation, producing a thousand cells that had exactly the same pattern of gene activity and that were entirely functionally equivalent to one another, and then spontaneously and suddenly switched to an intricate spatial pattern of differential gene activity.
What nematodes do is exactly what other animals do (with a bit more hardwired efficiency), defining pattern gradually by natural molecular mechanisms at the cellular level.
But evolution can’t select for an embryo!
This part had me staring open-mouthed at the screen. This was such an idiotic argument. Paul Nelson says,
Natural selection can’t select a future function. It can only select features that are advantageous already.
Say what? This is true if you’re talking about generations in a population; I can’t undergo selection for a trait that does me no good, but might be advantageous in my grandchildren.
But Nelson is applying this rule to stages of development. No, no, no.
With this rule, you’d have to argue that human testes could not have evolved, because boys have no use for them until they reach puberty. Fetuses could not develop a digestive tract, since they don’t eat, and would only be able to form a mouth and gut and anus when they were confronted with a breast.
This is absolutely nuts. In this case, the unit of selection is the viable individual capable of reproduction; all selection sees is whether this organism manages to replicate itself. There is selection against individuals that fail to produce testes or a gut, and of course there can be selection for individuals who can produce elaborations of their form that enhance the reliability of replication.
He calls himself a philosopher of biology
. How can a philosopher make such a stupid error?
Read the goddamned literature. I know they’re deeply attached to their own ignorance, but man, read some papers. It’s not as if the evolution of the development of nematodes is something no one ever thought of, until the clowns at the Discovery Institute called it to our attention. There’s a lively discussion of how it came to be, and it’s easy to find real papers in science journals that discuss it.
Here’s one by Bob Goldstein, On the Evolution of Early Development in the Nematoda (pdf). I can understand how Nelson would miss it, it’s not as if any of the obvious keywords are in the title, or something. One thing that’s very important to point out in any evolutionary analysis of nematode development is that the pattern he’s pointing out is specific to C. elegans — other nematodes exhibit variations. Here, for instance, are the four-cell stages of 35 different species of nematode.
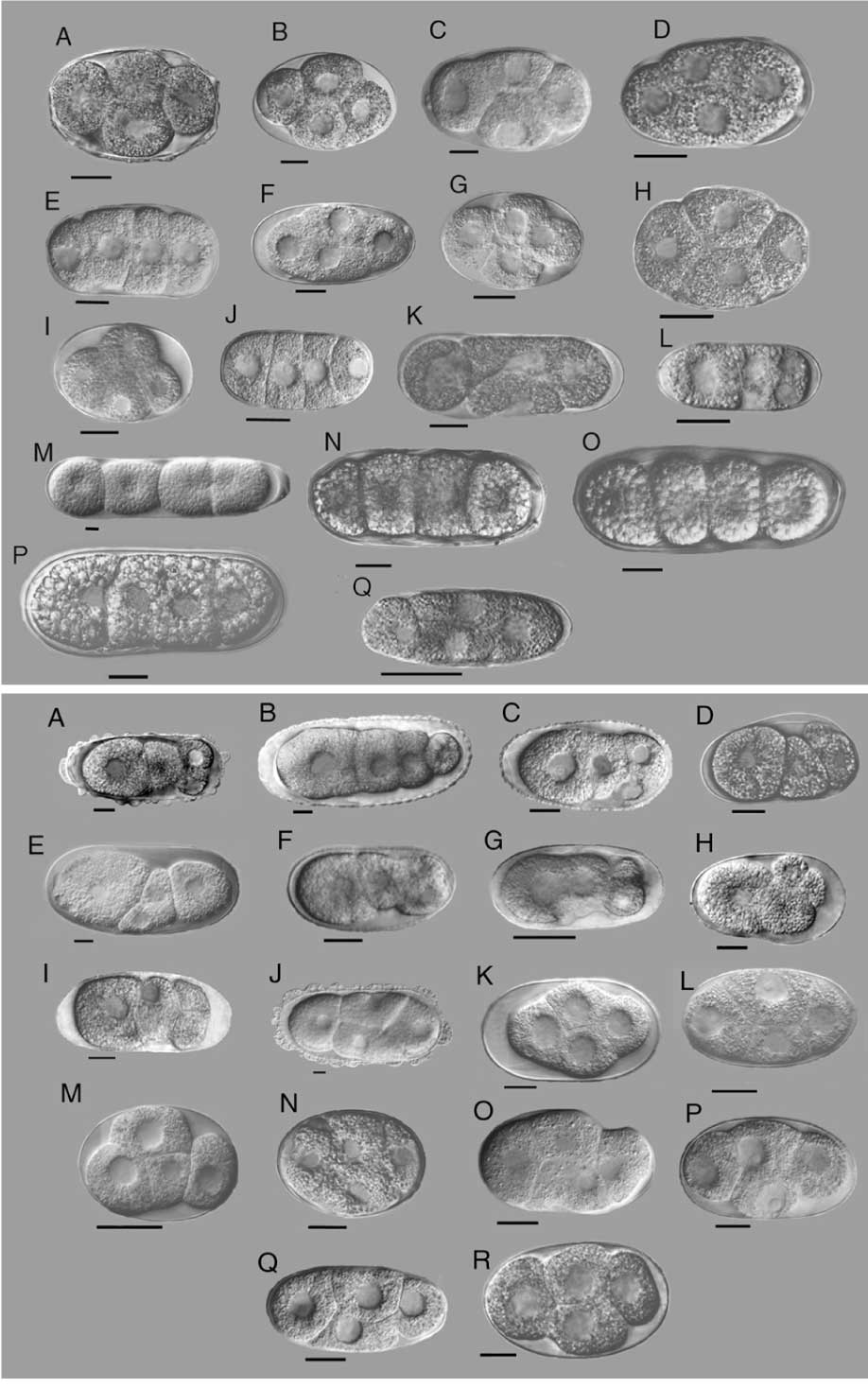
Orientation of cells at the four-cell stage in 35 species. Families and genera are, for (a) A-J, Rhabditina; A, Teratorhabditis; B, Bunonema; C, Cruznema; D, Mesorhabditis; E, Diploscapter; F, Rhabditella; G, Rhabditis; H, Caenorhabditis; I, PS1010; J, Protorhabditis. K, Aphelenchidae (Aphelenchus). L, Aphelenchoididae (Aphelenchoides). M-P, Tylenchina; M, Belonolaimidae (Belonolaimus); N, Heteroderidae (Meloidogyne); O-P, Pratylenchidae (Pratylenchus and Nacobbus). Q , Teratocephalidae (Teratocephalus). (b) A-I, Cephalobidae; A, Chiloplacus; B, Zeldia; C, Cephalobus; D, Acrobeloides; E. Acrobeles; F, Eucephalobus; G, Pseudoacrobeles; H, Nothacrobeles; I, Cervidellus. J-L, Diplogastrina; J, Diplenteron; K, Aduncospiculum; L, Pristionchus; M, Goodeyus. N-Q , Panagrolaimidae; N, Halicephalobus; O, Panagrellus; P, Panagrobelus; Q , Panagrolaimus. R, Brevibuccidae (Plectonchus). All scale bars are 20 mm.
Nelson is basically making the poker hand fallacy, with I’ve dealt with before when dealing with Behe’s nonsense.
Yet his argument for this dramatic conclusion is not only weak, it’s wrong. I could, for instance, correctly argue that the odds of getting a straight flush dealt to you in a 5 card poker hand is about 1 in 6×104; we could calculate this with probability theory, and we could also deal lots of poker hands and determine it empirically. No one’s going to argue with that part of the math.
But now, if I were to define a Straight Flush Complexity Cluster (SFCC) parameter and wave it around and claim that “no hand of the same complexity as a straight flush has been dealt by chance in the last ten years of poker games here in town,” that players can only possibly win one hand in 60,000, or worse, that no one has won a poker hand without cheating and stacking the deck, you’d know I was crazy. But that is basically Behe’s entire argument — he claims to have found the “edge of evolution,” and that it is much sharper and steeper and more impassable than anyone but a creationist could believe.
Creationists love to point to specific, complicated things and declare that that detailed configuration could not possibly have arisen by mere chance. But the thing is that when you look at all of the possibilities, that there are many other alternative ways of achieving similar results, the odds begin to go down and down and down, often reaching the point of absolute certainty.
Here’s Nelson’s gob-smackingly stupid and arrogant conclusion.
Neo-Darwinian theory has never explained the origin of animal development for this very reason. Today a range of incompatible evolutionary hypotheses is on offer, but none has solved the puzzle. As I say in the article, it is the biological community’s prior insistence on a materialistic explanation that keeps them stumped and stuck.
The evidence points unmistakably to a cause with foresight; i.e., a cause with a mind. I hope this helps.
After blathering on and on about the remarkable level of understanding we have of how nematode early development works, Paul Nelson pathetically announces that scientists are stumped and stuck
, and offers his dim, wrong, silly misconceptions to help
. I can guarantee you that no sensible nematode biologist is going to pay the slightest attention to that drivel from the Discovery Institute, except perhaps to laugh.
As a palate cleanser, anyone interested in more detail on the processes involved from some real scientists can go read about how asymmetric cell divisions contribute to axis formation. Unlike Nelson, who is stupefied by his own ignorance, the authors there can see clearly how evolutionary thinking helps to understand these events.
As discussed above, the mechanisms of cellular polarity and asymmetric division that determine the AP axis in C. elegans have clear parallels in other organisms. Likewise, the specification of the other axes of the C. elegans embryo, as well as that of particular cell fates, utilize highly conserved signaling modules, including the Wnt and Notch signaling pathways (see Wnt signaling, Notch signaling in the C. elegans embryo and LIN-12/Notch signaling in C. elegans). Thus, just as the study of the early C. elegans embryo provided key insight into developmental processes, studies in other systems inform the work in C. elegans to give a broader picture of the mechanisms that control development.
Goldstein B (2001) On the evolution of early development in the Nematoda. Philos Trans R Soc Lond B Biol Sci 356(1414):1521-31.
Murakami T, Hijikata T, Matsukawa M, Ishikawa H, Yorifuji H. (2006) Zebrafish protocadherin 10 is involved in paraxial mesoderm development and somitogenesis. Dev Dyn 235(2):506-14.

Part of the problem is that creationists can basically copy and paste a textbook and add “god did it” at the end. This is how people can take Behe seriously, not a lot of it is wrong, because most of it is just basic biochemistry.
Bad faith aside, what they miss is “it didn’t have to be this”. Anything that works will do, anything that doesn’t work is dead. So there are potentially Vast (Dennett’s usage of the word) numbers of potential solutions, of which we only observe the minute number that actually happened.
Who designed the designer. Work on that one PN. It might take you a millennia to figure out the truth, which starts with not accepting presuppositionally your imaginary deity poofed into existence “as is”.
Also note how creationists keep confusing “evolution” for “mere chance”.
When ever I hear about the long odds of something happening, I am always reminded of scenario in one of the old video games, where the undercover cop had to beat the casino in order to get the money to try a drug deal.
Of course, wining several slot machine pulls in a row was nigh impossible. The best strategy was to save the game prior to gambling, then if you won some money, save the game anew. So you were starting a new position, with more money, but needed less wins in fairly quick secession to reach your goal. Often four or five intermediate saves were enough. Evolution operates in the same fashion, with each generation, if there is any small advantage, saving the previous wins and by chance trying things to be in a better position in future generations. Eventually you get to what you see now through small, incremental, changes. Which when random selection is taken into account, the chance is close to 1.
Why not? I thought menopause was the usual example of precisely a trait that can be selected for even though it does *you* no good, it just helps your grandchildren.
Nerd: I love the analogy of evolution looking essentially like save-scumming in hindsight.
It’s all written down in the cosmic background radiation!
…And, actually, if one believed in Laplace’s intelligence […] assez vaste (later misrepresented as “Laplace’s demon”)—which is probably incompatible with quantum mechanics, but that’s hardly the only unbelievable thing about it—then a sufficiently precise knowledge of that radiation would be “the cosmic blueprint”, not just for the nematode but for the entire cosmos:
I don’t think Laplace was even a Deist (although, given his great ability to adapt whatever political philosophy would further his survival during, before, and after the Revoluton, he may also have applied the same tactics to theology), just an over-enthusiastic believer in the power of analysis (computation). But Charles Babbage (of the difference engine) was certainly some kind of orthodox Christian, and he would (I think) have been happy—had he known about the cosmic background radiation (or any other time-slice of the universe)—to endorse the notion that it was a “blueprint.”
Actually, though, I suppose that’s all carbon paper, not actually blueprints. Or something.
Um… no.
Apparently not covered in 9th grade biology, and I didn’t take college bio – just read a lot of random things which were mostly genpop articles about current research and mostly not from the turn of the 20th century.
Do I lose points? I’ll just do my Baymax-standing-in-the-corner impression over here.
Nerd @5
The creationists think evolution plays craps while it’s actually playing yahtzee.
Crip Dyke @9
Is such ignorance possible? :P
Did their designer make those little demodex mites that live in your face pores until their asses explode?
Menyambal @ 12:
Yes.
Face-pore ass-explosions are hella big yuks beyond the pearly gates.
“What exactly are you using as a criterion to differentiate evolved from designed?”
The Bible, duh!
Remember that Nelson’s got this bizarre linear view of evolution which starts with a single cell creature, which evolves into a creature with a few cells, which evolves into one with a few more cells, and so on until you reach the 1031 cells in the nematode today. It wouldn’t surprise me at all if Nelson thought that the creature at the 150 cell stage in this process had to be like a modern nematode at the 150 cell stage of development.
OK, OK: Here’s a summary of E.G. Conklin.
Yes.
I understand.
golkarian@1 — you’re giving Behe far too much credit. Even before his first famous neo-creationist book, Darwin’s Black Box, he was spouting major errors derived from his ignorance of the fields he was trying to dismiss. You can read Kenneth R Miller’s contemporaneous review of Darwin’s Black Box for examples.
The point is not that Behe made mistakes — everyone does, including Darwin and Einstein — but that his mistakes were (i) drawn from ignorance of a field he was “overthrowing” but not familiar with, (ii) drawn from significant ignorance of his own field of biochemistry, (iii) completely obvious to someone who doesn’t have his particular axe to grind (e.g. his “front-loading” argument makes zero sense in any biological theory, not just evolution), (iv) could have been easily avoided simply by asking some of his colleagues for critiques, (v) most importantly, the errors lead to critical failures with regards to his own theories, by which I mean he made these basic errors only because acknowledging them would catastrophically demolish his version of ID.
So, no, he doesn’t get a “mostly right on the biochemistry” pass from me. He doesn’t get brownie points for listing every complex bit of biochemistry he can think of only to use those examples in completely dishonest ways even within his field of biochemistry. That’s like saying Jenny McCarthy may be wrong when she links autism to vaccines, but at least she describes autism symptoms mostly accurately. Or that astrologers are at least right about the existence of stars.
Irreducible stupidity.
I wonder if these confused creationists can even figure out how to untie this math riddle. It’s a small demonstration of confused thinking in just a few short sentences.
just a small point: the scale bars are microns not mm.
Well as Anri says, that just shows the Designer* has a sense of humour! I mean, He’s**a laugh-a-minute kind of guy***: smallpox, polio, ebola, HIV, filariasis, Guinea worm, malaria – get the irreducible complexity of those parasitic life cycles!
* Or of course, Designers
** Or of course She’s, or Xi’s, or It’s or They’s *wink wink nudge nudge*
*** Or of course gal, non-gendered or non-binary person, computer, committee or collective intelligence cough*praisethelord*cough
When you’re stupid, EVERYTHING looks complex.
#20 brian – no, $49+$49+my$+mom’s$+dad’s$ = $101.
Where did the extra dollar come from?
No, wait a minute, now I’m ALL confused.
I have compared Paul Nelson’s use of words and phrases with those used by the blogger. Here is a list of words and phrases used by the blogger which are not based in science or objectivity but are obviously intended to discredit Paul Nelson by negative emotional response:
obsession
laughed at
pompously earnest
nitpick of an annoying detail
sloppy and shallow
cute trick
idiotic argument
absolutely nuts
stupid error
goddamned
clowns
nonsense
gob-smackingly stupid and arrogant conclusion
blathering on and on
pathetically announces
dim, wrong, silly misconceptions
drivel
stupefied by his own ignorance
Here are a couple of statements the blogger uses for the same purpose:
“I’ve got palm impressions in my forehead from smacking myself so many times while watching this terrible little video.”
“This statement is just wrong.”
Now here are all the similar usages I found in the words of Paul Nelson’s video:
.
.
.
Oh well, I couldn’t find a single one!
Thus it is not difficult to tell which one is being more credible and honest!
And the one thing missing from Paul Nelsons writing is credible scientific evidence from the peer reviewed scientific literature, properly presented and in context. Personal incredulity isn’t scientific. That is where bad thinking starts.
How an argument is presented has absolutely nothing to do with the truth of the argument. PN cannot prove his deity is real, and not a figment of his delusions. That is why he is mocking and shown to be a fool.
Joseph Morrow@25,
That’s because Paul Nelson is pretending – probably even to himself – to be a real scientist. So he adopts the gravitas he thinks appropriate to that role. Meanwhile real scientists, like PZ, don’t have the same need to “perform” being serious scientists: they have real credibility, real knowledge and real understanding, not the ersatz versions on display in that pathetic video.
Further to #27, Paul Nelson is like the complete duffer at golf or tennis, who buys all the snazzy gear and learns the technical terms, feels himself on a level with – or above – the winners of the “Masters” in their chosen sport, but continually swings and misses whenever he tries to connect with the ball.
@25 Joseph Morrow
If you are pointing out that PZ is sometimes rude, I’m pretty sure everyone in this forum already knew that.
It doesn’t change the fact that Paul Nelson is a buffoon. A polite buffoon, perhaps, but still a buffoon.
Paul Nelson also has a great schtick on Metamorphosis, available wherever fine creationist tripe is
soldpandered.BZZZZZ!! Wrong. Scientific facts are not affected by emotion or politeness, but thanks for playing.
If Paul Nelson or anyone else wants to avoid being insulted by a professional developmental biologist, he needs to stop making “scientific” claims about development he arrived at by his merely sitting in an armchair and declaring materialistic evolution impossible because his own two-bit intellect can’t imagine it happening. He needs to get off his butt, learn developmental biology as it is understood today, and then go spent thirty years in a lab collecting real evidence. Then, MAYBE he will be able to competently discredit some bit of the science. But probably not. Short of doing all that, he is simply exposing himself as a clown for gainsaying the informed opinions of his betters.
Joseph Morrow:
Speaker A: “With respect, good sir, the sky is fuschia.”
Speaker B: “You’re an idiot, the sky is blue, and here is a photograph that shows it.”
Who speaks with more credibility and honesty?
Sometimes I truly wonder how idiots like Joseph Morrow are able to breath independently. Must take up most of his brainpower.
Stupidest argument for creationism ever?
I’m not the first person to ask this question, by any means, but what kind of a monster actually consciously designs ebola? Or loa-loa worms, or ichneumon wasps? Or Lou Gehrig’s disease? Why are so many animals designed to survive by eating other animals alive? Why are so many people prone to addiction to substances which damage and eventually kill them? The list goes on and on.
The vast amount of gratuitous suffering in the world should make any reasonably intelligent person realize that if a creator exists, he is malevolent, incompetent, or both.
That must be the absurdest tone troll we ever had.
Fucking.
Insert word where desired.
@Joseph Morrow
You object to the use of
On what basis is it wrong to use that statement? Why should asserting that someone made an error be off limits?
Really, tone trolls – by definition – make clear that their priorities are more with dictating other people’s range of acceptable expression than they are with getting the best/right answers.
Usually, however, they try to deny or obfuscate that fact.
Congratulations, Joseph Morrow, on having the courage of your I’d-rather-everyone-be-polite-and-ignorant-than-have-feathers-ruffled-in-the-course-of-actually-learning-something convictions.
Didn’t creationists request — nay, demand — that scientists explain every little detail of how we got from Organism A to Organism B? Or how the differences between C, D, and E evolved? How can they turn around and say that’s not proof of evolutionary development in the past? Hmmph.
Do we know how phosphorylation of DNA occurs? Can this be described from an evolutionary process of random mutation and natural selection?