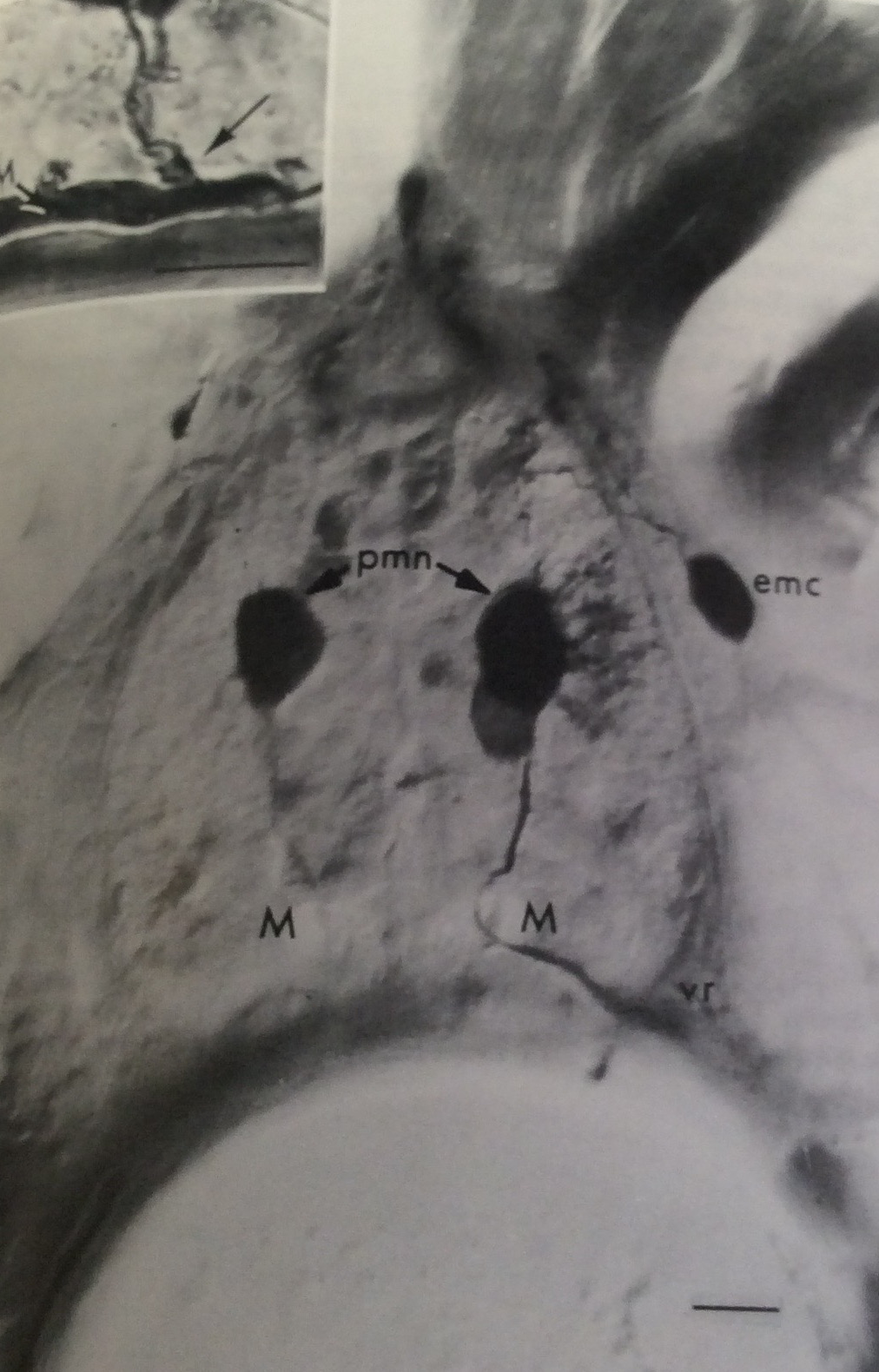
I know this is a horrible photo — I just snapped a picture of the journal hardcopy, which I own, instead of grabbing a PDF from the web, because it’s from 1985 and I’d have to pay to get a copy of my own paper — but this is what I was doing in grad school. I started as somebody who was interested in neurons and the nervous system, so what you’re seeing is a transverse section of the spinal cord of a zebrafish, with a couple of motoneurons labeled black with a tracer enzyme. I spent most of my time teasing apart how those cells grew and made connections.
But all the while there was this one prominent feature of the animal that kept trying to distract me. See that big clear white space below the spinal cord? That’s the notochord. It’s huge, a long transparent cylinder built like a stack of glass coins, running from the neighborhood of the hindbrain all the way back to the tip of the tail. The image from Stemple below is much clearer, since the notochord has been painted pink.
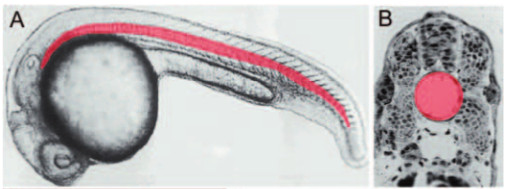
Its superficial function is obvious: it’s a springy rod that the muscles of the fish’s body act upon for swimming and escape behaviors. While I was just doing neural circuitry work, that was sufficient — it’s part of the motor apparatus. Neurons make muscles twitch which flexes the notochord and generates the back and forth motion that propels the animal through the water. Case solved.
Except…you can’t think of the notochord all by itself. Fish don’t flex vertically, they undulate laterally, and that implies auxiliary structures have to be present in order to constrain vertical movement. In particular, on top of the notochord is a specific part of the spinal cord, the floorplate, and beneath the notochord is another fibrous tissue, the hypochord. Together, these help constrain the movement of the animal for swimming.
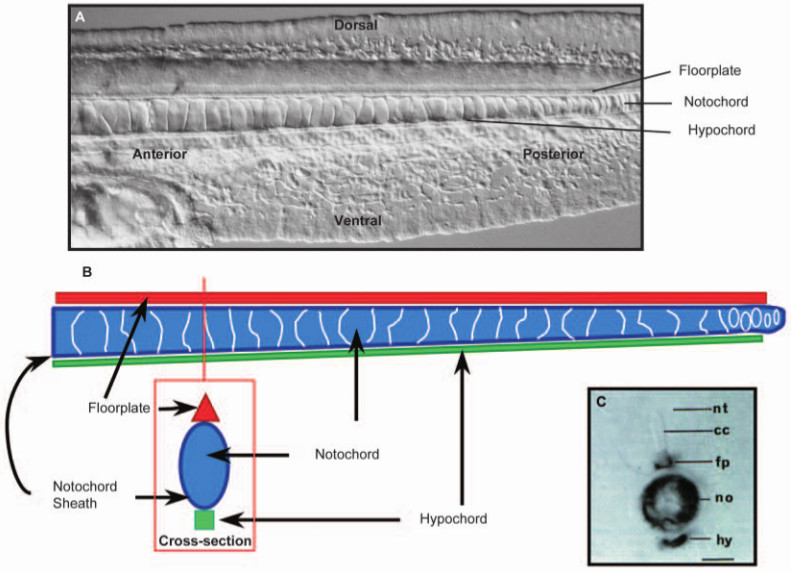
I can’t describe it any better than Stemple did, so I’m stealing this:
To understand notochord structure, it is helpful to consider the notochord as part of a mechanical system required for locomotion (Fig. 4). Zebrafish embryos, for example, are able to flip their tails within 1 day of fertilisation, and hatch and swim within 3 days. By analogy, the notochord is like a fire hose, possessing a strong but flexible sheath that can resist high hydrostatic pressures. Consider then a situation in which the fire hose is filled with water balloons, each pushing against the other and against the sheath. In such an arrangement, the fire hose would be elongated and stiff, but able to bend in any direction. Finally, with cables running along the top and bottom of the inflated fire hose, the cables would resist any upward or downward bending of the hose, and any force acting on the fire hose would deflect it laterally. For the zebrafish embryo, the equivalent of the fire hose is the peri-notochordal basement membrane and the water balloons are the vacuolated notochord cells. Running along the top of the notochord is the floor plate and along the bottom is the hypochord. Consistent with this structural role, both the floor plate and the hypochord express a variety of cartilage proteins, such as type II collagen. In addition, mutations in genes such as oep and cyclops, which lead to substantial loss of floor plate, also produce embryos with a profound downward curvature. Thus, although not yet directly established, it is possible that the floor plate and hypochord act as cables on the respective dorsal and ventral sides of the notochord.
But wait! There’s so much more. The notochord isn’t just a mechanical structure, it’s also a source of inductive signals that tell the spinal cord to form neurons. The first growth cones — the motile tip of a growing neuron — emerge where the spinal cord meets the notochord, and they creep along the surface of the notochord before charging off into the muscles. If I wanted to know how the nervous system builds that circuitry I was studying, I should look further into interactions with that long rod.
Also, I was beginning to appreciate the importance of the notochord to everything. In gastrulation, it’s those first cells that tuck in and migrate into the interior of the animal that are going to form axial mesoderm, which will eventually form the notochord. Oh, what a devious structure: it got me thinking about things other than the nervous system that develop, as well as appreciating how development is an integrative process that can’t actually be thoroughly understood by examining organs in isolation.
The floorplate is induced by the notochord — that is, chemical signals from the notochord instruct cells of the nervous system to differentiate into the floorplate. The notochord also instructs mesoderm to make the horizontal septum, the seam between dorsal and ventral blocks of muscle that runs the length of the animal. And then floorplate in turn signals to other cells in the spinal cord to differentiate into motoneurons, and interactions at the horizontal septum trigger formation of fast and slow muscles in the periphery. Basically, every thing I was studying was a product of negotiations in development between the notochord and nerves or muscle.
So I have to blame the notochord for my drift from neuroscientist to developmental biologist.
There was further wickedness. Notochords are kind of unique to chordates (with qualifications I’ll discuss below). All chordates have them, and they’re absolutely central to normal development of the organism, but — shock, horror — cephalopods don’t, and they develop just fine without one. This is actually the subject that first drew me into evolutionary biology, and as a nice side effect, also made me interested in cephalopods and other invertebrates.
Because, damn, the deep phylogeny of the notochord is a wicked mess. For a good, brief introduction to the evolutionary history of the notochord, I recommend a recent review by Annona et al.. Basically, in the early 19th century, Cuvier’s view that animals could be divided into four grand embranchements, groups that represented distinct body plans, prevailed. The world had the radiates (cnidarians and echinoderms), the articulates (arthropods
and annelids), the molluscs, and the vertebrates, and each had unique features that meant one could not possibly have evolved into another. The vertebrates, for instance, had the notochord, which had no homolog in any other animal, therefore imagining some transitional form between vertebrates and any other of the embranchements was pure folly.
Well, that wasn’t going to stand. Even in Cuvier’s day, Etienne Geoffroy St. Hilaire was speculating about ways arthropods and vertebrates could transform into one another. Then one breach was discovered in Cuvier’s scheme: tunicates, which Cuvier had placed in the molluscan embranchement, were discovered to have a notochord in their larval stages, which required a taxonomic scramble to put them in the same phylum with vertebrates, as chordates. During the nineteenth and early twentieth century, there was an almost incomprehensible dash to discover the homolog of the notochord in all kinds of invertebrates. Members of every other embranchement were at some point proposed as the ancestor to the vertebrates: obscure structures, like the stomochord of enterepneusts, were proposed as the precursor to the notochord on the basis of appearance or position in relation to other tissues. Looking back on it, it’s almost embarassing how hard scientists struggled to fit weak data and limited character knowledge to an evolutionary model. Just for completeness’ sake, here are some diagrams of the various schemes people contrived to generate a vertebrate from an invertebrate.
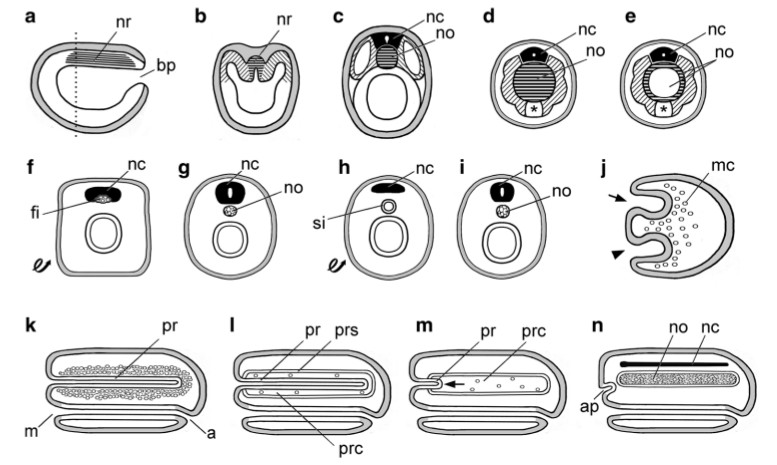
Notochords in phylogeny: invertebrate chordates, annelids, and nemerteans. a Midsagittal section of early neurula of amphioxus with blastopore (bp) and notochord rudiment (nr, horizontal hatching). b Cross section at level indicated by dotted line in a; notochord rudiment (nr) and somites indicated, respectively, by horizontal and diagonal hatching. c Cross section of later neurula of amphioxus showing notochord (no) and dorsal nerve cord (nc); somites indicated by diagonal hatching. d, e Early and late larvae, respectively, of an ascidian tunicate. Cross sections through the tail showing nerve cord (nc); muscles (diagonally hatched) and notochord (no); asterisk indicates endodermal strand (discovered by Seeliger). f, g Inverted annelid scenario (after Semper); following inversion (f looped arrow), fibers (fi) associated with the nerve cord (nc) are precursors of the notochord (no) in g. h, i Variant annelid theory (after Ehlers); annelid after inversion (h, looped arrow), the position of the siphon (si) corresponds to the vertebrate notochord (no) in i. j–n Nemertean scenario (after Hubrecht); gastrula (j) has a first invagination (arrowhead) for gut and a second invagination (arrow) for the proboscis, while mesenchyme cells (mc) ingress into the blastocoel. Subsequently k, a through gut forms from mouth (m) to anus (a), and mesenchyme cells condense around the proboscis (pr). l Schizocoely produces a proboscis coelom (prc) and a proboscis sheath (prs). The arrow in m indicates the proboscis (pr) pulling out of the proboscis coelom (prc), leaving behind a few mesenchyme cells. In n, the mesenchyme cells in the proboscis coelom have extensively proliferated to form the notochord (no); the remains of the proboscis have become the anterior pituitary (ap), while the dorsal nerve cord (nc) has formed by the dorsal migration and fusion of the lateral nerve cords.
Don’t think about it too hard. You’ll go mad.
The definitive answer is not going to be determined by embryological morphology. The notochord is a structure that is strongly tied into the development of many tissues, so what we need to look at is gene regulatory networks — are there homologous patterns of gene activity in any of the hypothesized invertebrate homologs? And the answer so far is…no. In particular, one of the defining genes expressed in the vertebrate notochord is a transcription factor called Brachyury, also known as T in mice. None of the putative homologs express Brachyury, which suggests that any other regulatory genes are going to be irrelevant, as far as notochord fate goes.
That’s why I told you not to think too hard about that complicated diagram above. A lot of old models are being rendered obsolete by new molecular data.
That leaves us with an extremely interesting idea, that the notochord actually was a completely novel innovation in chordate evolution. Some peculiar population of Precambrian worms acquired a mutation that, for instance, coupled Brachyury expression to a band of early endomesoderm, which in turn caused vacuolation of the roof of the archenteron, pushing out a subpopulation of cells to form a tail, and it was good, and they thrived with this new springy tail.
You might have some reservations about that, if you’re concerned about where these genes came from. The thing is, Brachyury is really old, and is found in all of Cuvier’s embranchements. Its initial role seems to have been in defining the location of the blastopore, which led to further roles in specifying the body axis, and then, in vertebrates, was drafted into specific jobs regulating mesoderm.
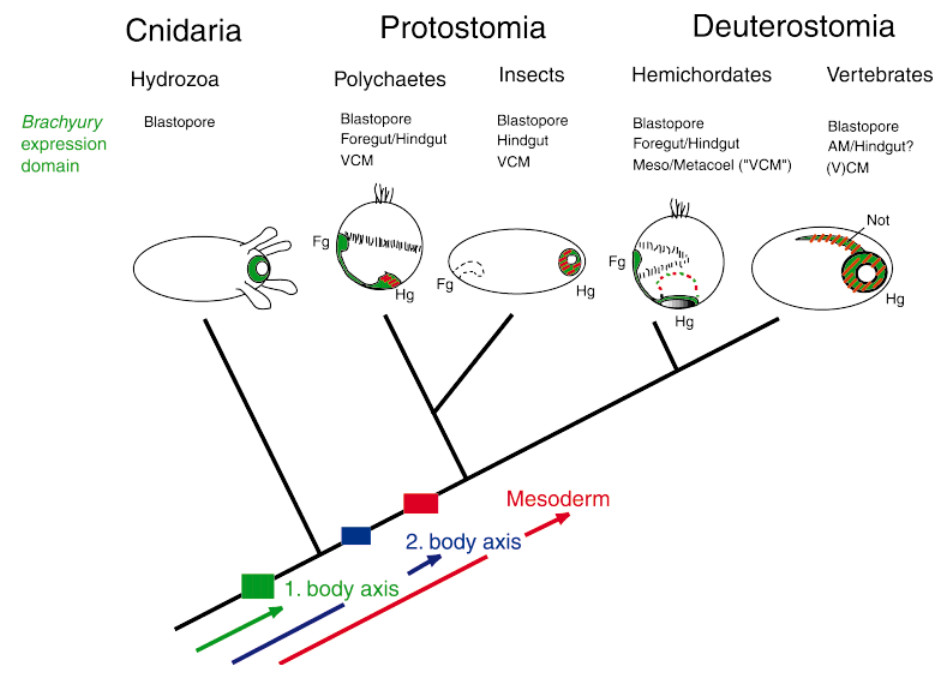
Scenario of the role of Brachyury during key events in animal evolution: emergence of the first body axis, the second body axis and the third germ layer, the mesoderm. Arrows intend to indicate that molecules implicated in these transitions were present before in a different context and that these transitions were brought about by inventing and recruiting developmental control genes to specific body regions or functions. These events might have occurred independently for the three major steps or, more likely, they may have been linked to and facilitate each other. Drawings represent gastrulae or post-gastrula larvae of different organisms. Brachyury expression is indicated in blue, presumptive mesoderm that overlaps with Brachyury expression regions in red hatched lines. Orientation is indicated by foregut (Fg) and hindgut (Hg). Other abbreviations: VCM, visceral cauddal mesoderm; AM, axial mesoderm.
So what was involved was not crafting a whole new assortment of functions, but wiring an existing gene into a network, where it then generated novel tissues. This is exactly why it’s not at all surprising that such different organisms as annelid worms, arthropods, and vertebrates share so many similar genes, yet have such different morphologies: what matters is how the building blocks of the genome interact with one another, so we can get different forms by changing switches.
Annona G, Holland ND, D’Aniello S (2015) Evolution of the notochord. Evodevo 6:30.
Stemple DL (2005) Structure and function of the notochord: an essential organ for chordate development. Development 132(11):2503-12.
Technau U (2001) Brachyury, the blastopore and the evolution of the mesoderm. Bioessays 23(9):788-94.

I loved this!
When I practiced pathology, one of my favorite stump-the-residents tumors was chordoma, with its physaliferous cells (with vacuolated cytoplasm as you describe above). And notochords have intrigued every biology student, since forever.
Thanks.
And chordomas express brachyury strongly!
“And chordomas express brachyury strongly!”
But politely, I assume, in Minnesota.
The diagram leaves out echinoderms, also deuterostomes. Shouldn’t their development be informative regarding this question? They are more closely related to the chordates than are the annelids and arthropods.
Yeah, and echinoderms don’t have a notochord, or anything like it…which further suggests that it is a vertebrate novelty.
Brachyury is older than old. It’s been found in some unicellular organisms including some fungi.
Excellent! I hope your students appreciate the broad integration of history, morphology, development, and evolution.
A quibble, however. I thought hemichordates had a sort-of notochord, and that tunicate larvae do too. Since the tree seems to be settling down to be:
((Echinoderms,Hemichordates),(Amphioxus,(Tunicates,Vertebrates)))
this would suggest that the notochord predates all of these, unless one can argue that the hemichordate “notochord” is a convergence, in which case our notochord would have arisen before amphioxus split off from us tunicates-and-vertebrates. Either way, it then would definitely not have evolved with the vertebrates.
OK, looked up hemichordates on Wikipedia, and they argue that the hemichordate “stomochord” is not a notochord but convergent. So on that view the notochord evolves by the common ancestor of amphioxus, tunicates and us vertebrates.
This is so fascinating.
I would like to understand more about morphogenesis, could anyone suggest me any resource on the specific topic?
Tunicates and amphioxus do have a notochord, clearly.
The stomochord homology is messy. It doesn’t associate with any muscles, which makes it weird, but Annona et al. say this:
So the expected network of molecules isn’t there, even if there are some similarities in signaling.
I once tried to play a notochord, but my pinky wouldn’t reach that elusive high note.
#7: Oh, right, I see where I wrote wrongly: the notochord would have been a novelty that emerged in the chordates, not the vertebrates.
I love it when you Science!