It’s my first completely free day of Christmas break! Grades are all submitted, nothing is hanging over my head, but I still got up at 5:30am and needed to do something, so I learned about spider gastrulation.
This was a disgraceful gap in my knowledge — I’ve worked on insects and on vertebrates, and am fairly familiar with gastrulation in those kinds of organisms, but one thing I did know is that there’s a lot of variation in the details of gastrulation, so every new clade seems to exhibit some novel way of tucking cells in to the early embryo. Spiders do it in a cool way.
There are fundamentals that don’t differ that much, though. Every animal has an early proliferative phase, the blastula, in which cells divide and divide and divide to generate a mass of cells, usually organized as a sheet. There are difference here, too: some extremely well-organized animals, like Drosophila only divide nuclei, making what is called a syncytial blastoderm, deferring the business of building membranes around them and creating separate cells until later. Other animals, like us, carry out full cellular divisions, so one cell becomes two cells, two becomes four, etc.
Others do something in between, carrying out nuclear and cellular divisions where it’s practical, but only partial membrane separations in other places. The chick embryo is a perfect example: the embryo proper is only a small disc on the surface of one spot on the gigantic yolk, and while some cells separate off from the yolk cell, other nuclei only have partial membranous separations — it would be energetically impractical to cleave the whole yolk into multiple cells.
So what do spiders do? They’re like the chick. Nuclei divide and migrate to a small disc on the yolk cell where they form a cellular blastoderm. Before any creationists get excited and think the similarity of spiders and bird blastulae as some kind of violation of common descent, no it’s not — it seems to be a mechanical outcome of different amounts of yolk in the egg. Lots of yolk = large cell, hard to divide, you get incomplete divisions. Less yolk = smaller cell and it’s more feasible to carry out full divisions.
But the question here is about gastrulation. Gastrulation is a process that takes a single-layered sheet of cells, and invaginates or folds in a subset of cells to produce the canonical three-layered animal embryo, with one layer committed to form gut stuff (the endoderm), another to form muscles, bones, blood (the mesoderm), and a third to form skin and nervous tissue (ectoderm). This is also important because the movement of one kind of cell past another kind of cell during migration creates novel interactions that allow multiple combinations of cell types. For instance, ectoderm makes skin and nervous tissue — what informs the layer about which one to make is the location of migrating mesoderm. As invaginating mesoderm slides past ectoderm, it instructs the ectoderm to develop into the nervous system, and where no mesoderm is present, the ectoderm defaults to making skin.
Beyond those generalities, there are significant differences. Amphibians have a single discrete blastopore, or entry point — cells stream towards that point and curl inwards to form mesoderm and endoderm. The first cells at that point form the organizer, a tissue that induces the central axis of the animal by, in part, signaling ectoderm to make nervous tissue.
Meanwhile, chick embryos have a long seam where ingression takes place. Drosophila…hey, flies also make a long seam! Again, don’t get misled by superficial similarities — it’s the molecular interactions that matter, and the mechanical arrangement of the tissues seems to be evolutionarily plastic. Other differences that can get embryologists quite worked up is whether the cells invaginate as a coherent sheet, or whether they delaminate and ingress as individual cells, or some combination of the two. Different clades seem to have differences here as well. What matters is that somehow, some cells will get into the interior of the embryo to set up these multiple layers and start signaling each other to make more complex patterns of cells and tissues and organs.
And this is where my curiosity about spider gastrulation is satisfied. Spider embryos have a discrete blastopore (sort of like frogs!), where cells are internalized to form a deeper mass of endomesoderm that migrates in one direction to define the spider body axis, and spreads out to form the deeper tissues of the animal.
There is also some terminology to learn. The germ band is what the segmented body axis is called; it’s the same term used in other arthropods, like Drosophila. The cumulus is the spider organizer (like in frogs), a cluster of cells that defines where the axis will form. This diagram might help.
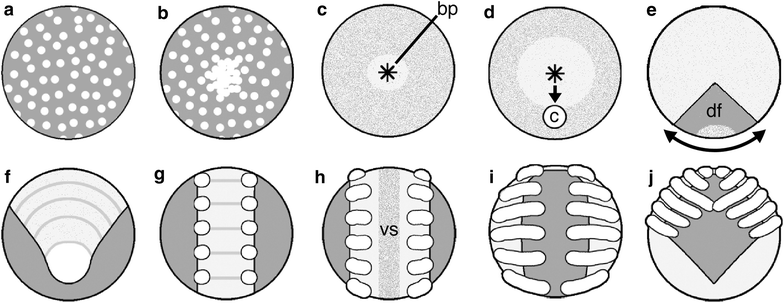
Canonical model of spider development, external view. a Blastoderm forms. b Blastoderm cells proliferate and migrate to one hemisphere to form a germ disc. c Primitive plate forms by internalization at a central blastopore. Asterisk marks blastopore (bp). d Cumulus (c) originates in the deep layer near blastopore and migrates radially to the prospective dorso-posterior side of the germ disc. e Germ disc cells move away circumferentially from the cumulus endpoint. This movement forms the germ band; the thinned area is termed the dorsal field (df). f Segmentation becomes apparent in the germ band. g Limb primordia appear on the anterior six segments (the prosoma). h The germ band splits along the mid-sagittal plane to form the ventral sulcus (vs). i The two halves of the germ band move laterally around the yolk, a process called inversion. j The prosoma condenses on the dorso-anterior side of the embryo and a sheet of cells surrounds the yolk.
If that’s not clear enough, watch the time lapse movies of spider development. They’re beautiful. Very leggy.
Note that the diagram above is labeled the canonical model of spider development. That’s because the paper I took it from is doing a comparative analysis of variation in gastrulation in different spider species, and finding differences. For instance, not all internalization occurs at the blastopore, but in some species cells will ‘leak in’ from various places around the rim. Delamination and ingression can occur in unexpected (by the standard model) places.
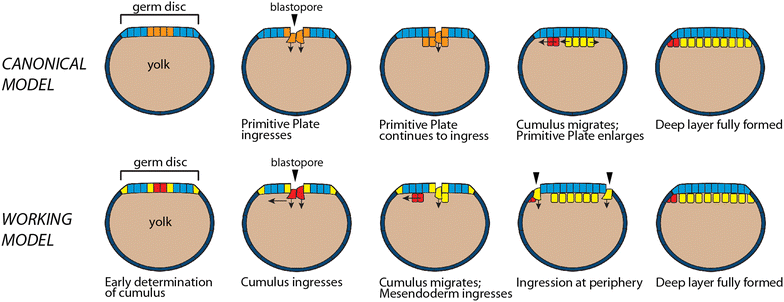
Proposed revisions to the canonical model of spider gastrulation. Diagrammatic midsagittal sections. Ectoderm is blue. Top row: Canonical model. A mixed population of cumulus cells and deep layer mesendoderm cells (orange) enters through the central blastopore. The cumulus (red) then differentiates and migrates towards the prospective dorsal side as the primitive plate (yellow) enlarges. Gastrulation ends when a deep layer has formed under the entire germ disc and signaling from the cumulus has broken radial symmetry. Bottom row: Our working model. The cumulus mesenchyme cells (red) internalize first, as a group, through the central blastopore. As the cumulus migrates, additional mesendoderm (yellow) cells internalize at two distinct locations: through the central blastopore and at the rim of the germ disc or, in species lacking a germ disc, a region several cells away from the central blastopore
These variations do not surprise me. Any time you do that evo-devo thing and look at development in different species, you’ll find a core ruled by a general principle and lots of differences in the detail. In the case of gastrulation, the idea is to get some cells into the interior to form new layers and for them to start talking to each other. In some organisms, the cells form orderly sheets and lines and march in coherently, in others it’s more like a mob pouring in any old way. It’s like the difference between driving in Canada and Italy: the same general, idealized traffic rules may be in place, but the implementation varies, with one being a more chaotic experience than the other. But they all get to their destination somehow.
Edgar A, Bates C, Larkin K, Black S (2015) Gastrulation occurs in multiple phases at two distinct sites in Latrodectus and Cheiracanthium spiders. Evodevo 6:33. doi: 10.1186/s13227-015-0029-z.

Last time I looked up gastrulation I wondered about the accuracy of the usual picture for protostomes vs deuterostomes. For example, the protostome page on wikipedia shows invagination sort of like a lollipop wrapper on a lollipop, forming a blastopore where the lollipop stick would go. The blastopose becomes anus or mouth, and then a hole erupts out of the top of the lollipop to make the other digestive end.
But is that actually how it works most of the time? When I looked up human gastrulation, it looked more like a crepe being rolled up with the ends being anus and mouth. The blastopore location was unclear. I ask because PZ mentioned frogs have a “discrete” (well defined?) blastopore, so maybe that’s unusual. Anyone know?
(I was reading about gastrulation for its connection to the abortion debate :)
Your lollipop analogy works for frogs. Not so good for humans, which form a flattened bilayer and then, like chicks, have invagination occur along a groove that extends from posterior to anterior. Mammals and birds are actually very similar, once you get past the initial yolky vs. yolk less state.
Fish have a cellular blastoderm draped over a large round yolk cell, with invagination at one region on the margin. If you were to flatten a fish or frog embryo, there would be similarities with mammals and birds.
Could this be what the estimable Steve Harvey meant in describing the universe as beginning in “a gastrous ball”?
Yes. After the Big Bang, the universe gastrulated. We’re now in the neurula stage, with galaxies folding in to form a colossal tube. When the Pharyngula stage arrives, I’ll be declared emperor and the whole universe will establish muscular organs that will allow it to swim away under my control.
We’ll leave Harvey behind, with Jesus.
And then I was, dead certain that spider embryos looked like balloon animals !?
This post just goes to show that even people who are very smart always have more to learn. I consider myself to be intelligent (then again, I’m pretty sure most people consider themselves intelligent) and I have always been hungry to learn things. I got a fairly broad education in college, though I admittedly did not focus much on biology or the other ‘life’ sciences. I did well in chemistry and physics, got a mathematics minor, a CS bachelor’s degree, and I’ve been working professionally for almost a decade. At this point I can effortlessly model complex computer systems from a blank sheet and have the knowledge and experience to build them. I think I am not wrong to consider myself some degree of expert in my chosen field.
Then I read something like this, knowing that there are people in their second year of college (and possibly some very advanced high schoolers) who grasp this the way I do code. My reaction to this article is quite simple: I think I know what some of those words mean.
Thanks, PZ, for helping me stay grounded. Maybe if I read this a few more times I might actually start to understand it. At the moment, all I have is “spider fetuses grow differently from non-spider fetuses, but within an understood and expected realm of possibilities”.
Cool, thanks for explaining. It would have pleased me to go around saying that humans develop “asshole first” like in the deuterostome picture, but it seems our development is more nuanced!
ealloc, haven’t you heard? We start out as a ‘clot of blood’ first. Duh!