Journey to the Microcosmos produces outstanding videos of microscopic life, and they have featured the volvocine algae before. This new video is entirely focused on the volvocines and on the evolution of multicellularity, and it’s really good.
Journey to the Microcosmos produces outstanding videos of microscopic life, and they have featured the volvocine algae before. This new video is entirely focused on the volvocines and on the evolution of multicellularity, and it’s really good.
My postdoc makes fun of me for having a lousy memory. Not long ago she showed me a paper about microRNAs, and I said I hadn’t read it. She responded, “Yes you have; you blogged about it!” The other day we were discussing the use of antibiotics to prevent bacterial contamination, and I said I thought I might have done that at one time. She told me I had, it was ampicillin, and the concentration.
I’ve been blogging for nearly four years now, and I’ve published well over 400 posts. So I’ve learned that before I sink a bunch of time into writing a new blog post, it’s worth a quick search to make sure I’m not going to repeat myself. When a new paper from Takashi Nakada and colleagues popped up in my Google Scholar alerts, I didn’t immediately realize that I had already written about it. That post was mainly about a new analysis by Thomas Pröschold and colleagues, with the Nakada trees serving as a point of comparison. The new paper is worth its own post, though.
A group of researchers from Keio University have published a new analysis of evolutionary relationships among green algae in the order Volvocales. Takashi Nakada, Yudai Tsuchida, and Masaru Tomita inferred relationships using one nuclear gene and five chloroplast genes.
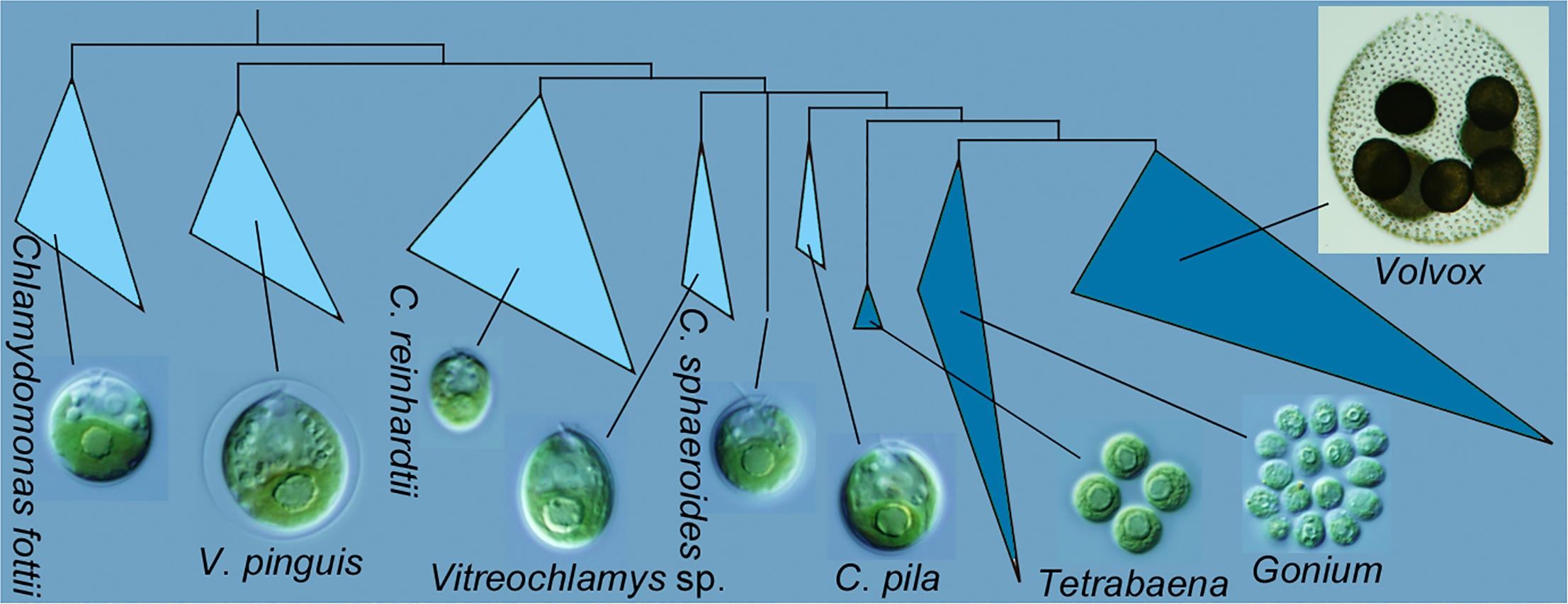
Graphical abstract from Nakada et al. 2019 showing Chlamydomonas pila as sister to the multicellular volvocine algae (Tetrabaena, Gonium, Volvox).
Previously, I focused on the monophyly of the multicellular volvocine algae, i.e. the Tetrabaenaceae, Goniaceae, and Volvocaceae (TGV). The multigene analysis shown above supports monophyly, although the support values for the critical node are not shown (meaning that the Bayesian posterior probability is <0.90 and the bootstrap proportions are <50%). Similarly, the new phylogeny doesn’t do much to resolve the backbone relationships within the Volvocaceae. There are differences from previous analyses that would be important if true, specifically in the positions of Volvox globator (the sole representative of Volvox section Volvox) and of Yamagishiella (which appears as part of an isogamous clade rather than sister to the anisogamous/oogamous Eudorina/Pleodorina/(most) Volvox clade). Neither of these differences is well supported, though, which is typical; most published phylogenies provide poor support for these relationships.

Figure 2 from Nakada et al. 2019. Bayesian phylogenetic tree of core-Reinhardtinia based on combined 18S-atpB-psaA-psaB-psbC-rbcL gene sequences. Corresponding posterior probabilities (≥0.90; left) and bootstrap proportions (≥50%) from maximum likelihood (middle) and neighbor-joining (right) analyses are shown next to the branches. Branch lengths and scale bars represent the expected number of nucleotide substitutions per site. Metaclades (MC; 1.00 posterior probabilities).
The main point of the new paper, though, is the close relationship between the multicellular volvocine algae and Chlamydomonas pila. The critical node for this relationship is is supported by a high Bayesian posterior probability (1.00) but crappy bootstrap values (55% for maximum likelihood and <50% for neighbor joining). The authors did do some analyses with fewer taxa to test this relationship, and those trees did have better support, but they also changed other relationships.
Correctly identifying the closest unicellular relative of the multicellular volvocine algae is critical for reconstructing the first steps in the transition to multicellular life. This is far from the first time that other species of Chlamydomonas and some of Vitreochlamys have been implicated. I’m not aware of any previous phylogeny that includes Chlamydomonas pila, but Chlamydomonas debaryana (for example) is usually closer when it is included.
I wouldn’t say that the evolutionary relationships in this group are fully settled at this point; the particulars vary among authors, depending on the gene(s) analyzed, and even depending on the method of phylogenetic inference. Even the monophyly of the multicellular species has been called into question, though I think it’s definitely too early to be confident in that conclusion. Right now it seems that Chlamydomonas pila is the best contender for the sister species to the multicellular clade, and almost certainly a closer relative to Volvox and co. than Chlamydomonas reinhardtii. As the authors point out, this makes C. pila a good candidate for whole-genome sequencing. The closer a relative to the multicellular group we can find, the better we can resolve which changes are specific to the multicellular clade.
Stable links:
Nakada, T., Tsuchida, Y. & Tomita, M. 2019. Improved taxon sampling and multigene phylogeny of unicellular chlamydomonads closely related to the colonial volvocalean lineage Tetrabaenaceae-Goniaceae-Volvocaceae (Volvocales, Chlorophyceae). Mol. Phylogenet. Evol. 130, 1–8. doi: 10.1016/j.ympev.2018.09.013

Plate from Pritchard 1834. Image from Google Books.
Andrew Pritchard’s 1834 book The Natural History of Animalcules includes several species he classifies as Volvox. Most of them were probably not Volvox, but his Volvox globator certainly was. His description of Volvox begins on page 39. A scanned version is available online at The Biodiversity Heritage Library, but I have used the slightly higher quality scan in Google Books for the plate above.
The animalcules belonging to this genus are of a globular form, and revolve in the water. Some of the species are so large as to be discerned by unassisted vision, while others are very diminutive. Ehrenberg has not demonstrated their digestive organization; but in a note to his table, conceives they ought to follow the monads. In this genus is included that beautiful animalcule, called the Volvox globator, which forms so interesting a spectacle in the Solar and Gas Microscopes.
Looking closely you can see the individual chloroplasts inside these Pandorina Morum colonies! Credit: Linden Gledhill pic.twitter.com/fk7LMdtneO
— Olympus Life Science (@OlympusLifeSci) November 29, 2017

Fig. 1 from Herron 2016. Examples of volvocine species. (A) Chlamydomonas reinhardtii, (B) Gonium pectorale, (C) Astrephomene gubernaculiferum, (D) Pandorina morum, (E) Volvulina compacta, (F) Platydorina caudata, (G) Yamagishiella unicocca, (H) Colemanosphaera charkowiensis, (I) Eudorina elegans, (J) Pleodorina starrii, (K) Volvox barberi, (L) Volvox ovalis, (M) Volvox gigas, (N) Volvox aureus, (O) Volvox carteri. Figure Credit for A and B: Deborah Shelton.
Pretty much what the title says: the meeting review from Volvox 2015 is online early at Molecular Ecology. That only took six months! This is the final, published version. Thanks for a great meeting, and thanks to everyone who read earlier drafts!
A new(ish) paper in National Science Review evaluates the evidence for various interpretations of Ediacaran microfossils from the Weng’an biota in South China (Xiao et al. 2014. The Weng’an biota and the Ediacaran radiation of multicellular eukaryotes. Natl. Sci. Rev., 1:498–520.). I recommend checking it out; it’s open access, and there’s a lot of interesting stuff in there that I’m not going to address.
These fossils are undoubtedly multicellular, probably eukaryotic, and extremely enigmatic. Their age (582-600 million years) means they could have important implications for the evolution of multicellularity, and their exceptional preservation in great numbers creates the potential for reconstructing their life cycles in great detail. Some of the Weng’an fossils have been interpreted as volvocine algae, an interpretation that I find highly unlikely.
Some of the Weng’an fossils are thought to represent red algae, and this would not be terribly surprising, since red algae have been around for at least 1.2 billion years. Others, for example the tubular fossils, are more problematic, with interpretations as diverse as cyanobacteria, eukaryotic algae, crinoids, and cnidarians.

Figure 8 from Xiao et al. 2014: Schematic diagram showing diagnostic features of the five recognized species of tubular microfossils in the Weng’an biota.

Fig. 62 from Weismann, A. 1904. The Evolution Theory. London: Edward Arnold. Pandorina morum; after Pringsheim. I, A young colony, consisting of 16 cells. II, Another colony, whose cells have reproduced daughter-colonies; all the cells uniformly alike. III, A young Volvox-colony; sz, somatic cells; kz, germ-cells.
Among the lower Algae there is a family, the Volvocinæ, in which the differentiation of the many-celled body on the principle of division of labour has just set in; in some genera it has been actually effected, though in the simplest way imaginable, and in others it has not yet begun.