As I’ve mentioned before, one of the things I’m doing in my Eco Devo class is to throw more of the burden of learning on the students. It would be too easy for me to just get up and lecture, telling them what they should know, and it is often hard for me to just shut up and let the students talk. I’ve split up the course so that Monday is when I start talking and dominate the classroom, Wednesday I ask the students to answer questions about Monday’s lecture and the book chapter, and on Fridays they’re given a paper to analyze.
This week’s paper is Morphological plasticity of the coral skeleton under CO2-driven seawater acidification by Tambutté and others. The context is that we’ve been talking about cellular physiology and development, and responses to environmental stresses, so I figured a primary research article about the effect of rising CO2 levels would be appropriate.
(Answer: more CO2 is not good for corals. Decreasing pH leads to a cnidarian version of osteoporosis.)
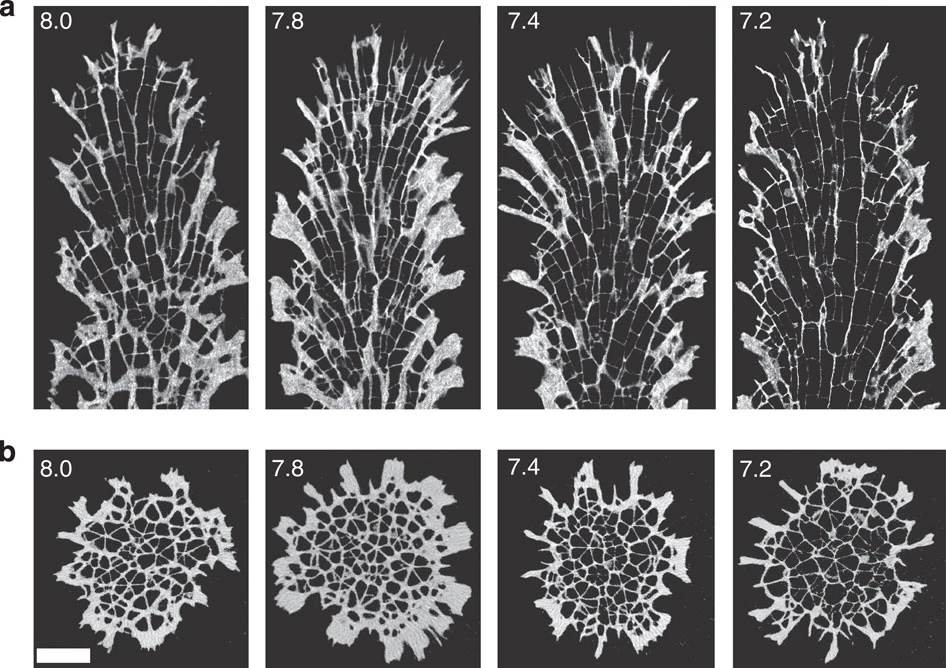
(a) Representative longitudinal sections; (b) transverse sections. pH treatment is indicated in the top left corner of each image. Scale bar, 1 mm.
















