I’m trying to raise money for the The Leukemia & Lymphoma Society, and I promised to do a few things if we reached certain goals. I said I’d write a post explaining what oncogenes are, while wearing a pirate hat, if we raised $2500. So here you go, arrr.

If you want more, go to my Light the Night fundraising page and throw money at it. I’ll write the next part when we hit $5000. Note that we’re also getting matching funds from the Todd Stiefel Foundation, so join in, it’s a good deal.
Cancer is not a creative, original disease; it has not been honed by ages of evolution to craft novel lines of attack on your body. Instead, it’s an opportunistic thief. Cancer misuses and perverts existing processes in your cells to send them out of control. Everything cancer does is simply the same thing your cells normally do, only amplified and unconstrained, driven by damage to the genes that would normally regulate their behavior.
Here’s a metaphor, a car with a dangerous defect. It has acquired a glitch in the accelerator so that every time you start it up, it immediately roars up to full speed, as if you’d floored the pedal. The problem hasn’t created anything new in the car, it’s just taken something you normally need to do, that is, regulate the speed of the machine, and stripped you of all ability to control it. That’s what an oncogene does; it is a gene that is normally involved in controlling the rate of cell proliferation, for instance, and a mutation has broken it in such a way that it now tells the cell to divide as rapidly as possible.
Now if you were driving down the freeway and suddenly your accelerator were stuck and you couldn’t slow it down, you’d have alternative strategies to stop (and so does the cell). You could hit the brakes or shift gears or turn off the ignition key. Cancers acquire another set of mutations that destroy the ability to shut off cell processes, analogous to breaking the brake pedal or snapping off the gear shift handle. These genes that can block the effects of out-of-control cell regulators are called tumor suppressors, and I’ll write about those at another time. Today I focus on oncogenes, regulators of the cell that must be damaged by mutation to produce an excessive response.
The first concern that comes to everyone’s mind is that you don’t want to have your cells running amuck — no one wants cancer. Just as you can do your best to maintain your car, you can also live sensibly — eat in moderation, avoid carcinogens or other behaviors that expose you to radiation, and get regular checkups — to reduce the likelihood of deleterious mutations. But they can happen anyway, through no fault of your own. Every time your cells divide, there is a very small chance of an error in replication that inserts a mutation into an oncogene. Just existing, even while doing everything exactly right to maximize your health, brings with it a base chance for a mutation. Given normal rates of cell division, every single one of you reading this is going to acquire about 20,000 DNA lesions today and every day. Almost every one of them will be patched up by DNA repair mechanisms (you have no idea how important DNA repair is to your continued health), but even so, one will occasionally slip through — over your lifetime, your cells will acquire an estimated 10,000 mutations. Live long enough, playing these odds, and cancer is essentially inevitable.
So cancer is fundamentally a chance process. There is no reason people get cancer, no purpose behind it, and everyone is susceptible. Some behaviors can increase the odds — smoking, failing to use sunblock — and you can also inherit genetic predispositions that increase the likelihood of acquiring a full set of mutations that lead to cancer, but ultimately, no one is at fault for cancer.
So what can go wrong? The diagram below is a simplified illustration of the various signaling networks in the cell. These are some of the pathways by which cells are told to regulate their behavior.

I’m going to focus on just the greenish box in the middle, the one labeled “Proliferation Circuits”, just to keep it simple. Think of that as the accelerator peddle for your cells. Sometimes your cells need to be encouraged to proliferate. For example, during childhood there are general signals to encourage stable patterns of growth, and during adolescence there may be novel hormonal signals to encourage new growth of selected populations of cells. If you’re injured, local growth factors are secreted to encourage cells to divide and repair the damage. So that’s what you’re seeing on the far left: growth factors and hormones can send a signal to the cell to give it permission to grow.
In order for a signal to be received, the cell has receptors on its surface that can bind to the growth factors and hormones. When a receptor binds to a signal, it changes to send a signal to other proteins (all those green circles) inside the cell. Think of it as like a doorbell; growth factor comes calling, presses the doorbell/receptor, the bell rings/sends a chemical signal into the interior, all the proteins get busy.
You can probably imagine how this system could get broken already. What if the receptor were damaged in such a way that it constantly sent a signal inwards, even if no growth factor were present (this is called becoming constitutively active)? What if one of the internal proteins were damaged in such a way to become constitutively active, so that it acted as if it were seeing the receptor as bound to a growth factor, even if it wasn’t? Now the cell is being constantly lied to by its sensory apparatus, and behaves as if it were being constantly told by the body to divide and divide and divide — it is on the road to being cancerous.
Let’s look a little bit closer at that pathway and give some of those green balls a name. This is the Ras-Raf signaling pathway.
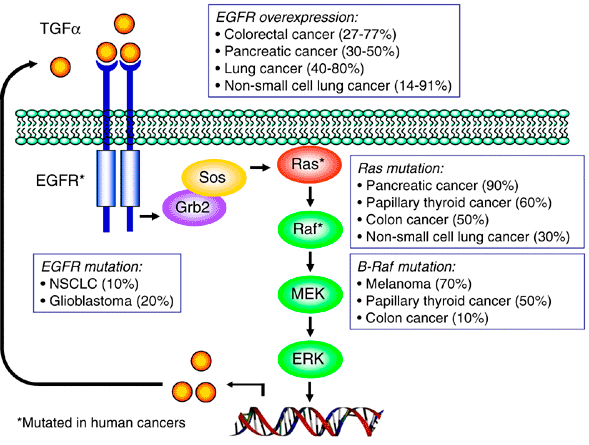
It’s a bit Rube-Goldbergian, but hey, that’s biology. In the top left corner you see something called EGFR, short for epidermal growth factor receptor. That’s our doorbell; the asterisk after the name in this diagram means that you see it mutated fairly frequently in human cancers. The box explains that it is seen to be damaged in 10-20% of certain kinds of cancers; when it’s mutated, it acts as if its signal, TGFα (Transforming Growth Factor Alpha) is always present. EGFR may also be perfectly normal, except that it is overexpressed, that is, present in a far greater number of copies on the cell surface, which makes the cell particularly sensitive to tiny quantities of growth factor. This happens even more frequently in many cancers.
The next steps in the pathway involve switching on Sos/Grb2, which activates Ras (look at the box; Ras is very commonly mutated in a great many cancers), which activates Raf, which activates MEK, which activates ERK, which promotes DNA replication. This is called a signaling cascade, and these kinds of sequences are all over the place in the cell. The advantages of this pattern is that many steps can be amplification steps, where one activated protein switches on many copies of the next protein in the sequence, and also each step is an opportunity for regulation.
Now be strong: I’m going to look even more closely at a key oncogene, Ras. Ras is important; it’s a central regulator of proliferative processes in the cell, and it is commonly one of the proteins made constitutively active in cancers.
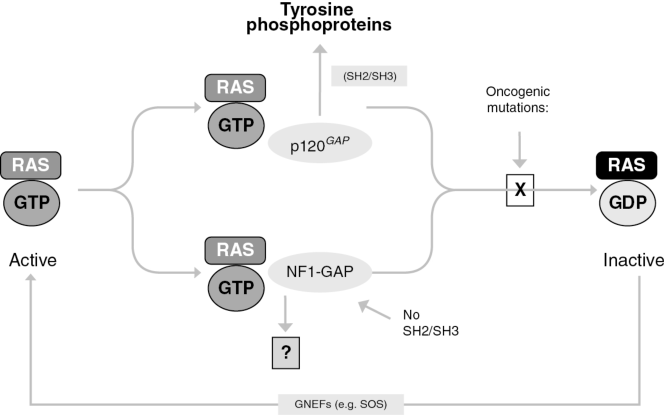
How is Ras switched on and off? In order to be active, Ras needs to be bound to a small molecule called GTP, guanine triphosphate, a base with three phosphate groups attached to it. It must be three; anything less, it isn’t active, so if it is bound to GDP, guanine diphosphate, it is inert. When it is bound to GTP, Ras changes shape to open up a binding site that can dock with the next protein in the chain, Raf, and activate it in turn.
Ras is switched on by the previous protein in the chain, SOS. SOS is a guanine exchange factor, and from the name, you can guess what it does: it swaps out the stultifying GDP molecule from Ras and replaces it with the energizing GTP molecule. Which allows it to bind with Raf, which then activates MEK, etc.
What switches Ras off? Ras is also an enzyme, specifically a GTPase — it cleaves its own GTP into a GDP. It turns itself off! What this means is that it acts as a kind of momentary switch. When a growth factor molecule arrives at the cell, Ras will respond by briefly initiating the cell division machinery, and then shutting itself down. It needs a sustained signal to keep cell division humming — a little triggering flicker of a random molecule bumping into the cell is not enough. This makes a lot of sense for cell cycle control. Ras is a reluctant activator, always hesitating and drawing back, and it needs constant prodding from external signals to keep doing its job.
Except when it’s mutated.
The most common cancer-inducing mutation in Ras is a single amino acid change in the 12th codon of its gene that greatly reduces the effectiveness of its GTPase activity. It binds GTP, becomes active, and then does not cleave the GTP — it clings to it instead. It switches from being a reluctant activator of cell division to instead being an avid, hyperactive activator — any transient signal, even a bit of noise, becomes an excuse to tell the cell to start dividing madly.
And that’s the beginning of a cancer, a rogue protein, made by an oncogene, that’s telling the normal, healthy cellular machinery to do its thing when it shouldn’t.
There’s much more to cancer than that, of course — cancer is more than just excessive cell division, and also the cell has many fail-safes, the tumor suppressor genes, that are supposed to put the brakes on when renegade proteins are going wild. But — oh, and this is cruel of me — to hear that part of the story, you’ll have to donate to my Light the Night fundraising page. When we get to $5000, I’ll tell you all about tumor suppressors.
Croce CM (2008) Oncogenes and cancer. N Engl J Med. 358(5):502-11.
Hanahan D, Weinberg RA (2011) Hallmarks of cancer: the next generation. Cell 144(5):646-74.
Hesketh R (2013) Introduction to Cancer Biology. Cambridge University Press.

That was excellent, clear and easy to understand. Thank you. Oh, and I like Pirate PZ better than Angel PZ.
This definitely sounds more plausible than cancer cells running some “ancient code” left by our evolutionary ancestors.
Good post!
PZ, this is awesome. Only 1/3rd of the way through so far, but loving it. My father is/was an oncologist (don’t know if he’s still working). B/c of his job, I learned a bunch about cancer from what he would tell me, and more from reading “Oncology Times” regularly for pretty much all of high school. But I don’t have a background in biology or medicine per se, and this is tremendously helpful. I’m already suddenly able to put things that I thought I understood into a new framework and realize that my previous understanding…wasn’t.
So for that, and for the Pirate Hat pic, huzzah!
I remember in the early 1980s as a graduate student in chemistry, happening to mention to the department head of biology about an interesting new concept I had read about: oncogenes. “Ridiculous! she commented. There’s no such thing as oncogenes – cancer is probably caused by a virus.” As your schematic shows, the science has come a long way since then!
Great post, PZ. I think I’ll forward this to some students.
IMHO, you make a convincing pirate. This may be the photo that reveals your true nature.
That actually happened to me once, when I was 19 and driving through Oklahoma on the way to join my husband who was in the Army in Texas. My solution was to turn off the ignition, which I believe is no longer possible on more modern cars. A guy came and showed me where the throttle cable had worked free of the nut that attached it to the accelerator, which is called that, even though what it actually does is prevent acceleration unless you push the pedal down and thus loosen the cable. At least it was so in late 50s era cars (hey, I was 19. It was an old car)
So oncogenes are what caused the nut to work off the cable. Okay. Got it.
PZ,
You make a great looking pirate!
I have autoimmune issues. My immune system is hyper and reacts to my own body as though my own body parts are the enemy. My salivary glands, eyes and other parts are dry due to Sjogren’s Syndrome and my muscles are affected by something, so I can hardly walk. Of course, the docs try to tell me that it’s all in my head.
Could any of this be explained by oncogenes?
Sorry if it’s a really dumb question – I’m a Latin teacher, not a scientist, Jim!
Yes! Very clear presentation, I loved the visuals (including the pirate PZ). Actually, I was hoping to know more about the role of cytokines, but I’ll shake my sad, little piggybank and see if I can help us all attain higher enlightenment at $5K for information on tumor suppressors.
magistratamarla:
Same here. I wish I knew what caused it, it’s a right pain in the arse.
Fantastic! I’ve been writing up lectures on lung cancer treatment for a few days without any and hadn’t had time to research this. Good to find out what I’ve been on about…
Finally got a chance to read the post. Great job explaining a complex subject.
I loved this. If I could have had you as a biology professor, I might have actually taken biology in college.
A friend of mine who has, shall we say, interesting views on diet posted this on Facebook the other day. Once I got to the “sugar feeds cancer!” bit my eyes rolled out of my head, but I don’t have anything remotely resembling the background to address the metabolic vs genetic theory. If anyone has the inclination and the time to read over it and give me something to use in discussing it, I would be very grateful. (If I were currently employed, I’d donate money in the hopes of getting to the $10,000 stretch goal, but money is pretty dang tight right now.)
Thanks for this, PZ! I’m eight years out from breast cancer treatment (lumpectomy, chemo, radiation) for a highly estrogen-receptive positive cancer (97% +). This truly helps explain exactly what was going on, and why everyone kept telling me that if I had to get a cancer, mine was one of the most treatable with a very high survival rate.
Being the control freak I am, I immediately studied up as best I could on my particular strain of cancer when I got my diagnosis and had laid out my plan of treatment, which DID NOT include chemo or radiation. Of course, you get 3 oncologists and a surgeon telling you that you’ll probably die if you don’t have the chemo and radiation and you tend to take their word for it, begrudgingly and resentfully, and are always positive that they were wrong and you would have been fine following your own course of action. But I digress badly.
Part of my reading led me to the work being done with dichloroacetic acid – DCA, the molecule which supposedly turns the mitochondria of a cell back on after it has been attacked by cancer, thus allowing the cell to die and taking the cancer with it. (And no, this was definitely not part of my plan for my own treatment since no trials had been done and its efficacy in treating different types of cancer was spotty, at best). I’m hoping to hear info that I can tie into this once you reach your next $ goal or two.
I’ll repeat what I and a fair few others have said before: came for the snark, stayed for the education.
Thank you, PZ. I suppose I’d better give you a few quid now.
A fascinating post that even a layman like your’s truly could understand.
Also, Pirate PZ is all kinds of awesome.
Yeah, and since he has access to well aged grog, he can supply the other ship with a couple of kegs as a friendly gesture, sail away, and come back and loot the ship without a shot being fired.
So I’ve been reading up on cancer and genetics a bit because my family seems awfully prone to cancer (grandfather, two out of three of his kids with four cancers between them, and a cousin). There do seem to be genetic mutations associated with cancer that can be heritable. Do they cause trouble by mucking with DNA repair, tumor suppressors, or the cell proliferation targets you talked about? Or all three?
Hmm…Dirty biological systems? The answer is most likely partially yes to all three.
Of course there’s a lot more to it, but this is a pretty succinct and accurate take-home message about what cancer is.
Thanks PZ! I’ve recommended this to my students, as the level of detail is just right for first-years.
I assume that’s 10,000 per cell? Could change to “each of your cells will aquire…” if so.
@Katiemarshall – sounds like a mutated tumor suppressor gene. I’m sure PZ will explain Knudsen’s two-hit hypothesis in the next installment…
All three. To my knowledge, tumor suppressors are the most common type of heritable cancer mutations. The reason for this may be that oncogenes tend to be gain-of-function mutations, and an active disturbance in the regulation of cell proliferation has a good chance of messing up embryogenesis and resulting in a nonviable fetus.
Most of the tumor suppressor mutations are loss-of-function mutations that tend to be autosomal recessive traits. The individual has one mutant copy of the gene and one good copy, and the good copy functions normally enough for embryogenesis to proceed to viable birth. The individual is at increased risk for developing cancers later in life because it only take one mutation to knock out their good copy of the tumor suppressor gene (whereas with other people they need two mutations, knocking out both copies of their tumor suppressor gene, in the same cell).
bognor#21:
Not counting neurons, your individual cells do not survive intact your entire lifetime; instead, they divide to pass acquired mutations to sister cells. 10k refers (not sure what he’s basing that number on though) to the estimate of somatic mutations, that are yours and yours alone. The estimated number passed on to you from your parents is about 60 or twice that depending on what papers you’ve read and how old your dad was at your conception.
PZ, +1
I thought it was more like 200 true mutations a day. Maybe 20000 includes all deletions, insertions etc.
I found this reference :
rorschach #25:
There’s a distinction between DNA lesions, the vast majority of which will be rapidly detected and corrected by DNA repair machinery, and true mutations, which are lesions that escape repair.
gillt #23:
Yep, ~60 new inherited mutations from your parents sounds about right, based on Nature Reviews Genetics 13:745 (2012). That says an average sperm or egg at age ~30 years has accumulated ~30 mutations (ignoring sex differences). Crudely extrapolating, a diploid (x2) somatic cell at age 60 years (x2) should have in the order of 30 x2 x2 = 120 mutations. That seems inconsistent with the estimate of 10,000 mutations per cell, unless gametes have vastly better DNA repair than somatic cells (which is possible, I guess).
The total number of somatic mutations that are yours alone in your entire body must exceed a quadrillion.
Every time your cells divide, there is a very small chance of an error in replication that inserts a mutation into an oncogene.
As cleverly illustrated by “pedal” later mutating into “peddle”…
Would you mind if I used this as a handout for clients? I am frequently asked to explain why a pet has developed cancer, and while the short version of “living in the world – it was nothing you did to your dog” is easy to break out in the exam room, it would be helpful to have something like this to send home with people to read later when they are less acutely distressed.
@bognor #26.
You shouldn’t ignore sex differences because there’s some evidence to suggest more mutations are inherited from the paternal side and this because a male’s germline undergoes more divisions.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3322360/
I know, but that wouldn’t affect a comparison between the sex-averaged number of de novo mutations inherited from parents, and the sex-averaged number of mutations in an adult’s somatic cells (which I presume is what PZ’s 10,000 figure means). Especially when the apparent discrepancy is in the order of 50-fold.
The higher mutation rate in non-germline (ie somatic) cells, on the other hand, is exactly what would explain the discrepancy, so thanks for the link.
The paper PZ Myers referenced above, “The Hallmarks of Cancer”? Buddhini Samarasinghe’s writing a series at Scientific American‘s guest blog explicating them. It’s an immensely fascinating series, because she’s targeting interested lay people. She explains the basics of molecular biology before she explains the paper.
And because of that her articles are a little clearer, I think. Certainly, I’m finding this article far easier to absorb after reading hers.
@8 and 10
Autoimmunity is not really mediated by oncogenes. There are a bunch of different ways for autoimmunity to occur, and some, but not all, share a somewhat similar outline (hijacking of something that should have been suppressed to become “on” all the time”), but the mechanisms are generally distinct.
@30. I’d still like to know how PZ calculated 10k mutations per generation. It’s a useful number to know.